A very long article on a very small croc…
Caption: a Koumpi montage, showing images by Mark Witton and from Barker et al. (2026).
If things go to plan – and that’s a big if – 2026 will see various announcements here relating to crocodylomorphs, that fantastic group of archosaurian reptiles that includes the crocodylians and their numerous extinct relatives. Today sees the first of those, and it relates to a publication, brand-new as of yesterday, of a super-detailed, extensive study focused on the small Wealden crocodyliform Koumpiodontosuchus aprosdokiti. Koumpi-what?
Yes, an animal that we’ll be calling Koumpi for short. Named in 2015 by Steve Sweetman and colleagues (Sweetman et al. 2015), Koumpi is represented by a small, near-complete skull (about 11 cm long) in addition to an atlas vertebra and ten osteoderms.

Caption: Koumpiodontosuchus had its time in the limelight back when it was first published in 2015. The original description (Sweetman et al. 2015) is open access at Acta Palaeontologica Polonica. The reconstruction here, created by Mark Witton and used in Sweetman et al. (2015), shows Koumpi manipulating a viviparid gastropod, carcharodontosaurian theropods and damselflies in the background. Image: Mark Witton.
When alive, the whole animal would have been about 80 cm long. Skeletal fusions and other feature show that it was near mature, and thus small as an adult. Its polysyllabic name really isn’t all that bad once you know how to break it down: it’s a suchus, and it’s koumpio (button) donto (toothed). Nevertheless, it’s a complex name to be sure, and one that might, just might, have been created as a semi-spiteful act of retaliation in response to the 2011 naming of another Wealden crocodyliform, namely Anteophthalmosuchus Salisbury & Naish, 2011. That name, also, is not so bad once you get to know it.
Yes, Koumpi is from the Wealden – that famous, highly productive Lower Cretaceous succession of sandstones, mudstones and siltstones – and specifically from the Wessex Formation of the Isle of Wight. Many of the Wealden dinosaurs I’ve worked on, including Eotyrannus, Thecocoelurus, Aristosuchus, the spinosaurids Ceratosuchops and Riparovenator and the big sauropod ‘Angloposeidon’, are from the Wessex Formation too and hence would have lived alongside Koumpi. Incidentally, the Koumpi holotype was found at Yaverland on the island’s south-east coast, as of course was the controversial Yaverlandia, a second specimen of which was recently described (Naish & Sweetman 2026). More on Yaverlandia is coming in time.
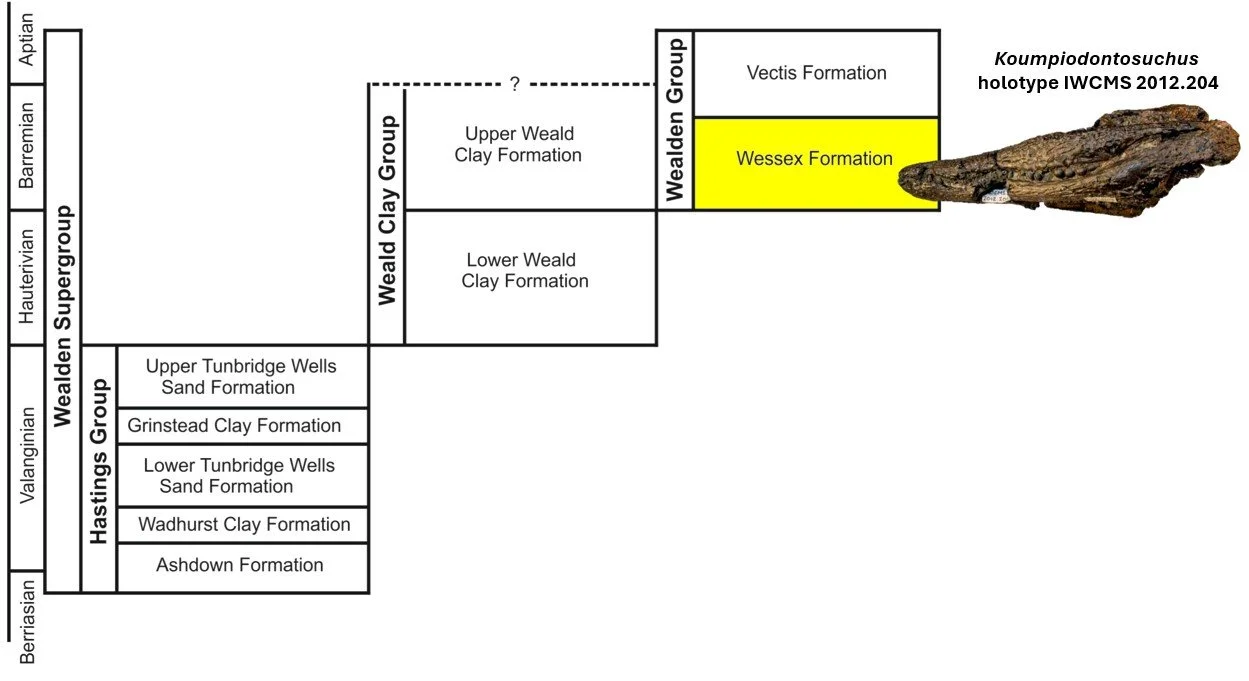
Caption: a simplified representation of Wealden Supergroup stratigraphy and terminology, again. The Wessex Formation, mostly Barremian in age, yields the greatest number of Wealden fossil reptiles, and Koumpi is among them. Image: Darren Naish; the Koumpi skull is from Sweetman et al. (2015).
Koumpi is heterodont, with caniniform anterior teeth and rounded, so-called tribodont, teeth at the rear of its jaws. The latter are suggestive of a diet that involved the crushing of shelled prey. As I keep saying whenever this comes up, such teeth do not indicate specialisation for durophagy (that is, a diet devoted to hard-shelled items), since living reptiles whose dentition includes at least some such teeth are generalists.
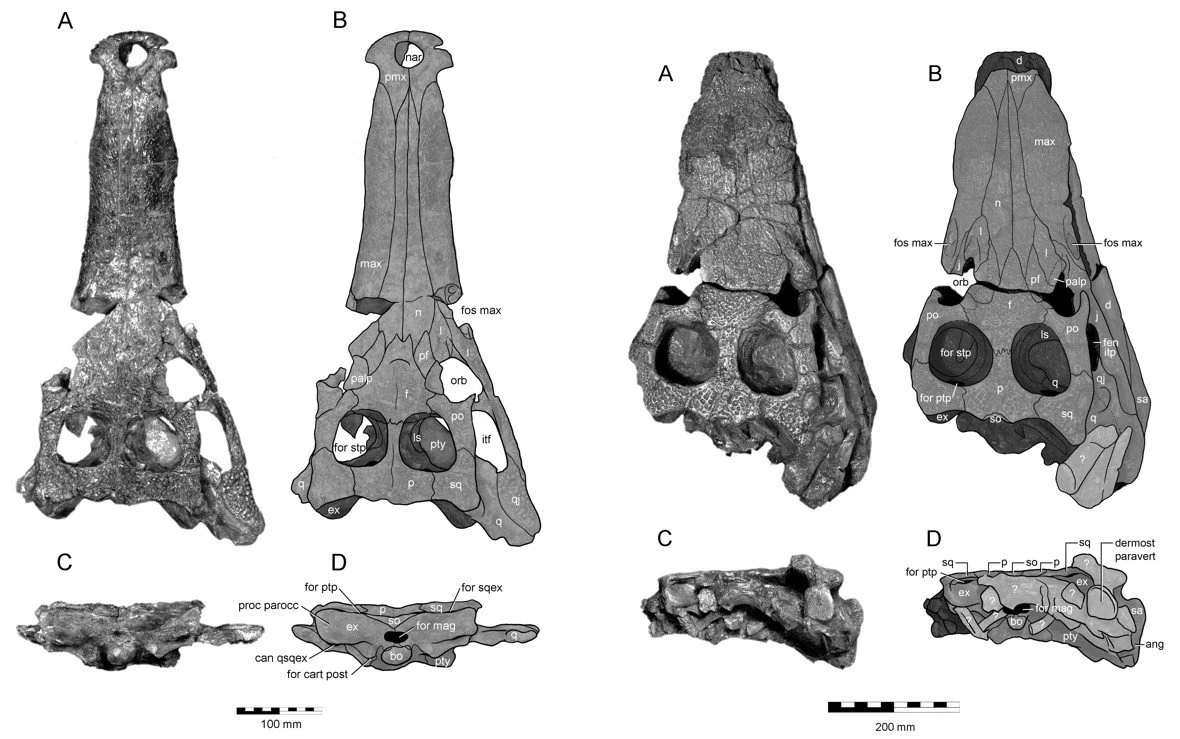
Caption: the Wealden crocodyliform assemblage includes an impressive number of goniopholidids, a group that is not the focus of interest here. These images show two of the best Wealden goniopholidids, Hulkepholis willetii at left, Anteophthalmosuchus hooleyi at right. Images from Salisbury & Naish (2011).
An important position in phylogeny. On the terminology in use for this group of animals, I’ll remind you that crocodylian (with a y, not an i) pertains only to the crown-group, Crocodylia. This is part of a much larger clade that includes numerous extinct groups, namely Crocodyliformes. This, in turn, is part of an even more inclusive clade – Crocodylomorpha – that includes lineages substantially different from crocodylians. A very unwise decision to disrupt and sabotage this taxonomy has been rebuffed (Brochu et al. 2009) and for good reason… for god’s sake don’t give up on it now!!
Within Crocodyliformes, crocodylians emerged close to the Jurassic-Cretaceous boundary from within Neosuchia, a group that includes lineages that, while undeniably crocodylian-like, lack the key specialisations of that group. These lineages – the goniopholidids, bernissartiids, susisuchids, hylaeochampsids and others – presage crocodylians in showing how their unusual palatal, vertebral and integumentary traits were assembled. Within Neosuchia, hylaeochampsids and some other groups are allied with crocodylians to form Eusuchia. I know that there are a lot of names to keep track of here, but what can I say… there are a lot of animals out there and the trees that depict their relationships are complex. There’s just no way round it.
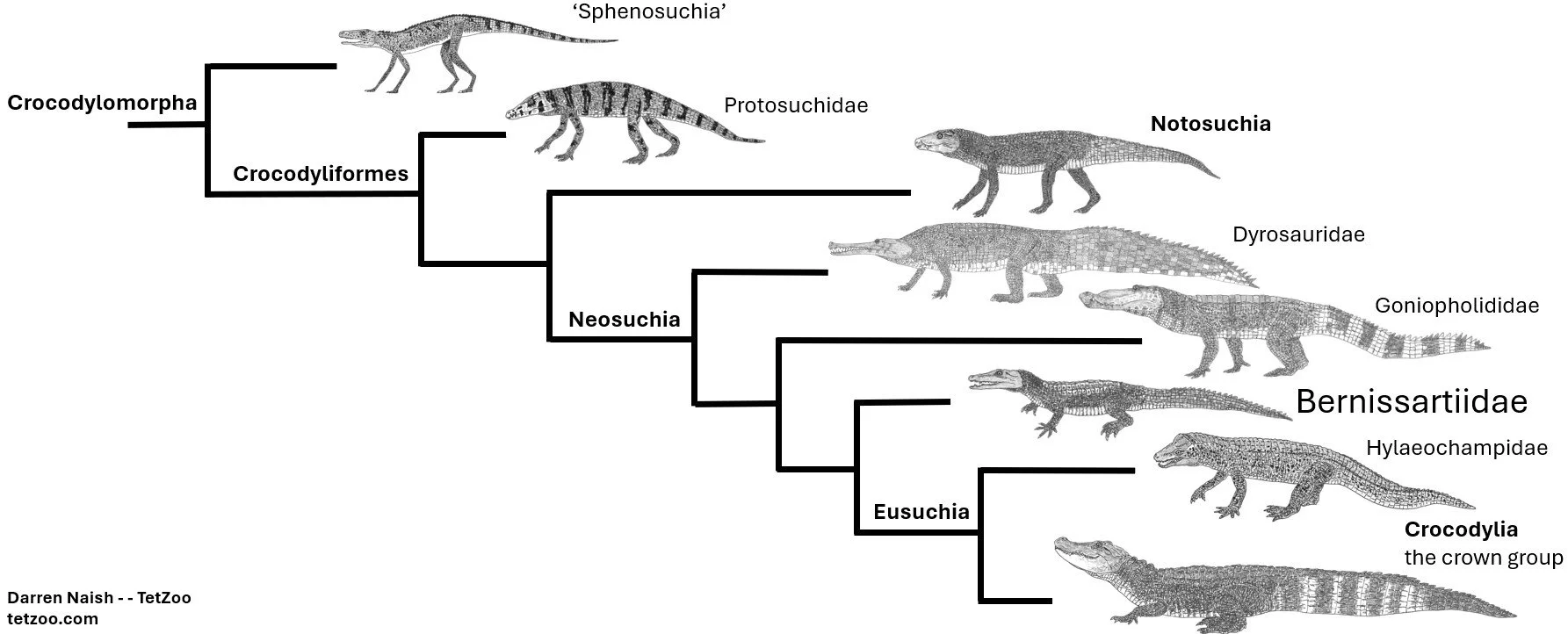
Caption: a substantially simplified crocodylomorph cladogram, showing the memberships of Crocodylomorpha, Crocodyliformes, Neosuchia and Eusuchia. Bernissartiidae, according to the bulk of studies, is outside of Eusuchia and close to eusuchian ancestry. Image: Darren Naish.
When first described, it was immediately evident that Koumpi had much in common with the famous Bernissartia, another small Wealden neosuchian (albeit known also from several Upper Jurassic European units as well). Bernissartia has long served as a pivotal taxon is discussions of crocodylian origins. Sweetman et al. (2015) confirmed this via phylogenetic analysis, finding Koumpi to be part of Bernissartiidae. That looked pretty solid to me. So it was surprising when Groh et al. (2020, 2022) found that Koumpi might be closer to hylaeochampsids or to paralligatorids, or that Koumpi and certain other Mesozoic neosuchians might be within Eusuchia (Martin et al. 2020, Groh et al. 2020, 2022).

Caption: the mounted Bernissartia fagesii lectotype specimen on show at the IRSNB (Insitut Royal des Sciences Naturelles de Belgique, Brussels). It’s small, at around 60 cm in length, and is behind glass so hard to photograph without reflections. Crocodyliforms of this sort have a biserial compliment of dorsal osteoderms, not a tetraserial one like that typical of crocodylians (albeit not present in all of them). Image: Darren Naish.
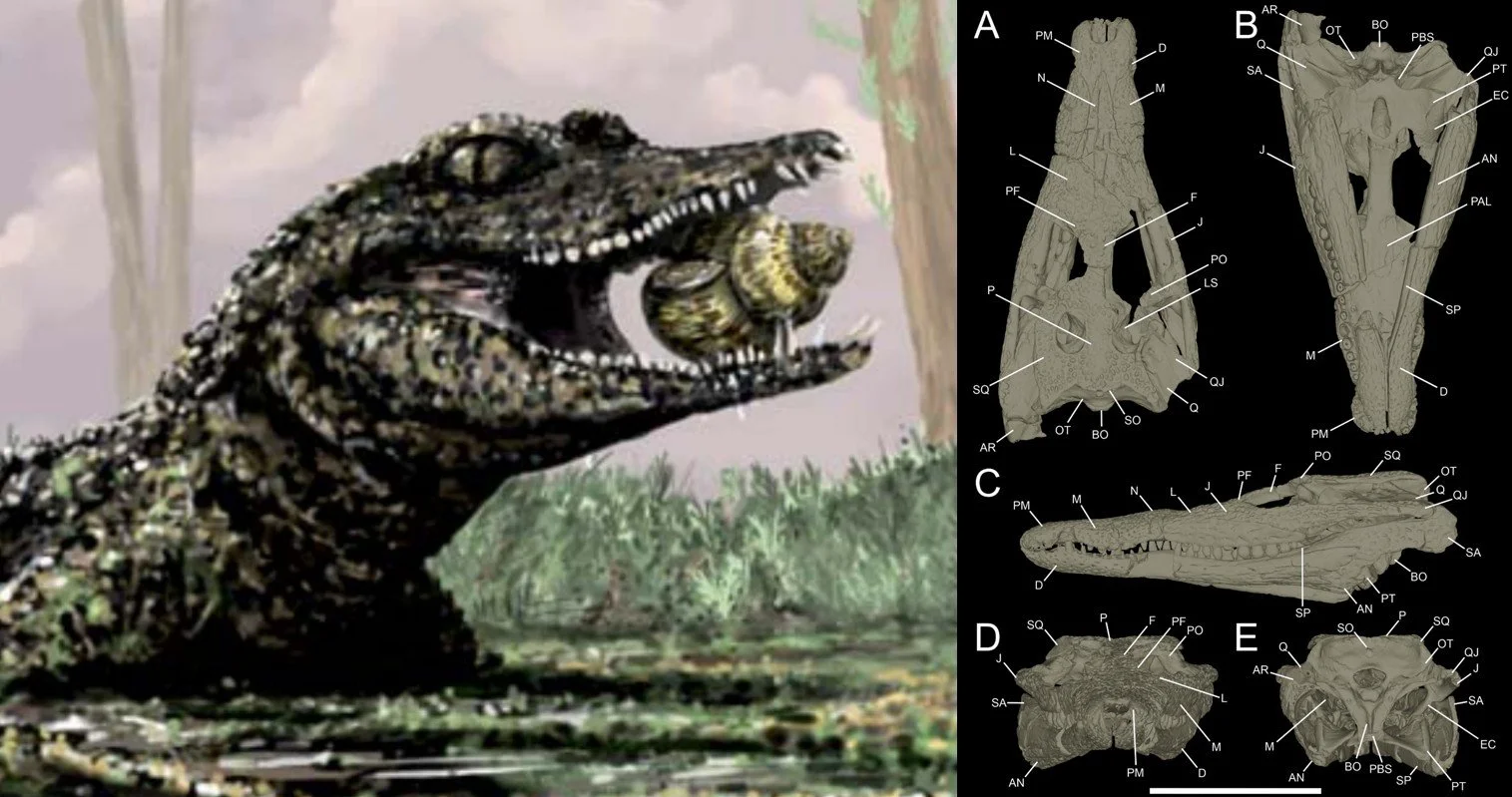
A comprehensive anatomical atlas. In view of the importance of bernissartiids to hypotheses of crocodyliform evolution and the excellent preservation and intact nature of the holotype Koumpi skull (not to mention its availability to our research group), I and a team of colleagues led by Chris Barker and Neil Gostling at the University of Southampton arranged a full microCT scan of the specimen and the production of a detailed description that substantially updates the initial one provided by Sweetman et al. (2015).
Caption: images from Barker et al. (2026), at left showing our retrodeformed, partly reconstructed model of the Koumpiodontosuchus cranium and mandible. At right, images showing blood vessels and nerves discovered during our analysis. Check the paper to see these images at full size. Images: Barker et al. (2026).
Our study – it’s open access – has just been published (Barker et al. 2026) and it forms a substantial anatomical atlas of the animal’s known anatomy. The data is so comprehensive and substantial that Koumpi now ranks as one of the most thoroughly described non-crocodylian neosuchians. Our other authors are Ethan Tulloch, Mark Young, Lai-Cheuk Leung, and Kathryn Rankin of the University of Southampton’s MuVIS X-Ray Imaging Centre (Barker et al. 2026).
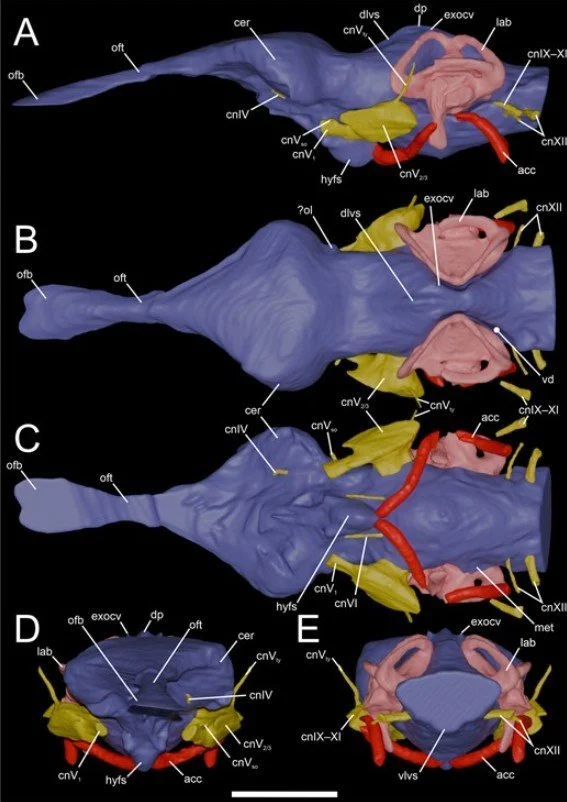
Caption: reconstructed brain, inner ear and associated blood vessels of Koumpi from Barker et al. (2026). The olfactory lobes are obvious and what might be the optic lobes can be detected too. The overall shape of the brain recalls that of immature crocodylians but we think that this might be paedomorphic trait, and other Koumpi brain traits are characteristic of adults. Images: Barker et al. (2026).
This new information enabled us to infer the hearing and neurosensory ranges of this animal, all indications being that it was similar in its abilities to extant crocodylians (Barker et al. 2026). This presumably means that there were ecological and behavioural similarities, which is what we’d predict in view of anatomical gestalt. We were also able to reconstruct the brain, the cranial nerves, the cerebral vasculature and the extremely complex paratympanic pneumatic system, this being a convoluted, multi-part arrangement of air-filled cavities and tubes connecting the middle ear and throat cavity (Barker et al. 2026).
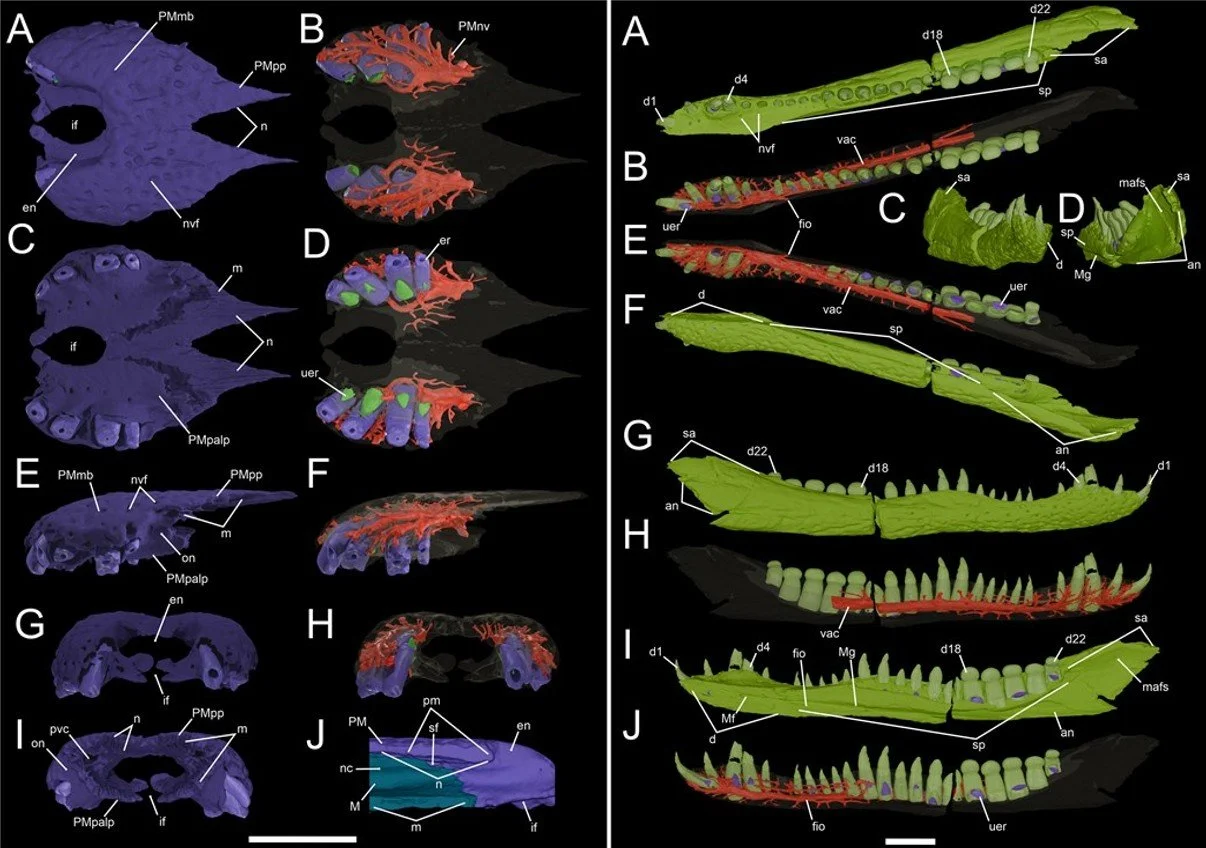
Caption: more of our excellent anatomical imagery of the Koumpi cranial material. At left, the conjoined premaxillae, some of the images showing (in red) the neurovascular canals. At right, the dentaries, showing (in red) the ventral alveolar canal, the tooth roots and more. Those tribodont posterior tooth crowns are especially obvious here. Images: Barker et al. (2026).
Phylogeny re-evaluated. We were also able to use our new anatomical data to look anew at the phylogenetic position of Koumpi. We evaluated those proposed affinities with hylaeochampsids and suggested placement within Eusuchia and did not find them supported (Barker et al. 2026). In contrast, we found strong support for the ‘conventional’ placement of Koumpi within Bernissartiidae, and for a placement of Bernissartiidae close to, but not within, Eusuchia (Barker et al. 2026).
Given that we incorporated a substantial amount of new anatomical data pertaining to the anatomy of the skull – over 240 new character states were coded for (Barker et al. 2026, supplementary data) – I’m going to make the argument that our assessment is much more data-rich than previous ones and therefore more likely to be correct. Koumpi is, once again, firmly back among the bernissartiids.
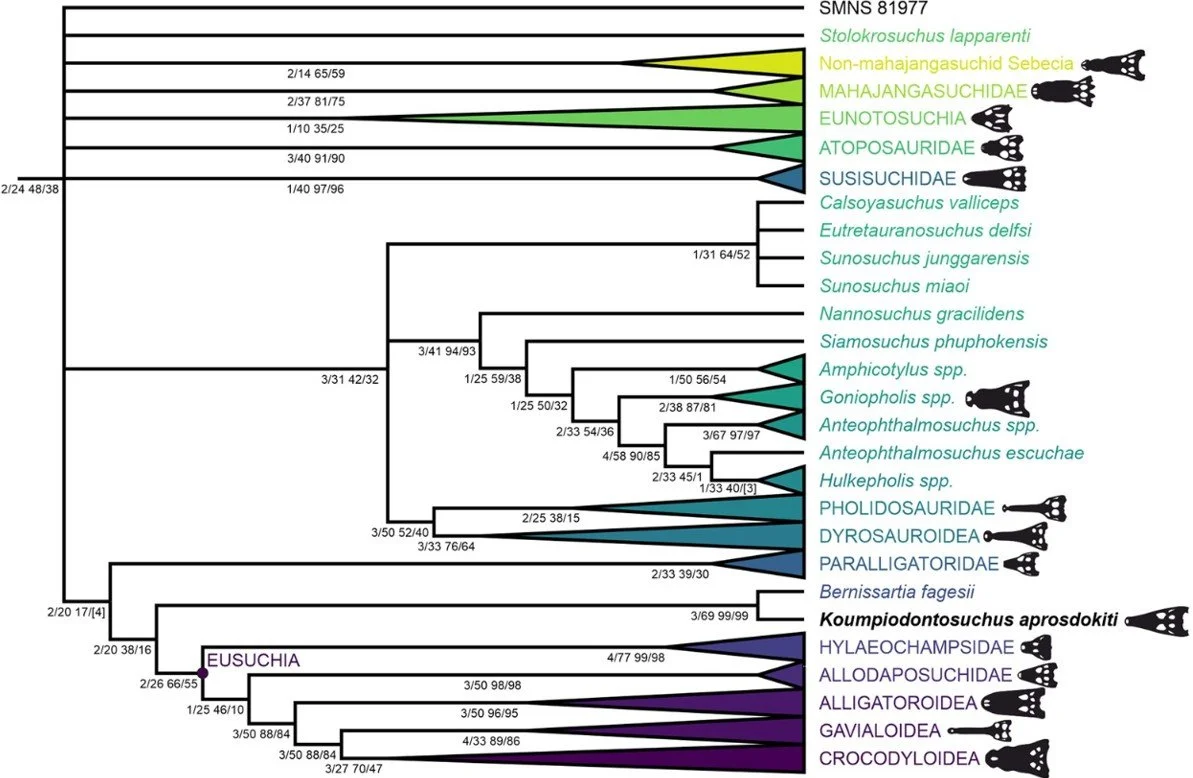
Caption: a strict consensus phylogeny resulting from our ‘equal weighing’ analysis, showing poorly resolved relationships among several lineages close to, and within, Neosuchia. The recovery of a clade that includes goniopholidids and tethysuchians (dyrosaurids and kin) is interesting, albeit not novel to our study. Note that Koumpi is a bernissartiid, and that bernissartiids are one step outside Eusuchia. Images: Barker et al. (2026).
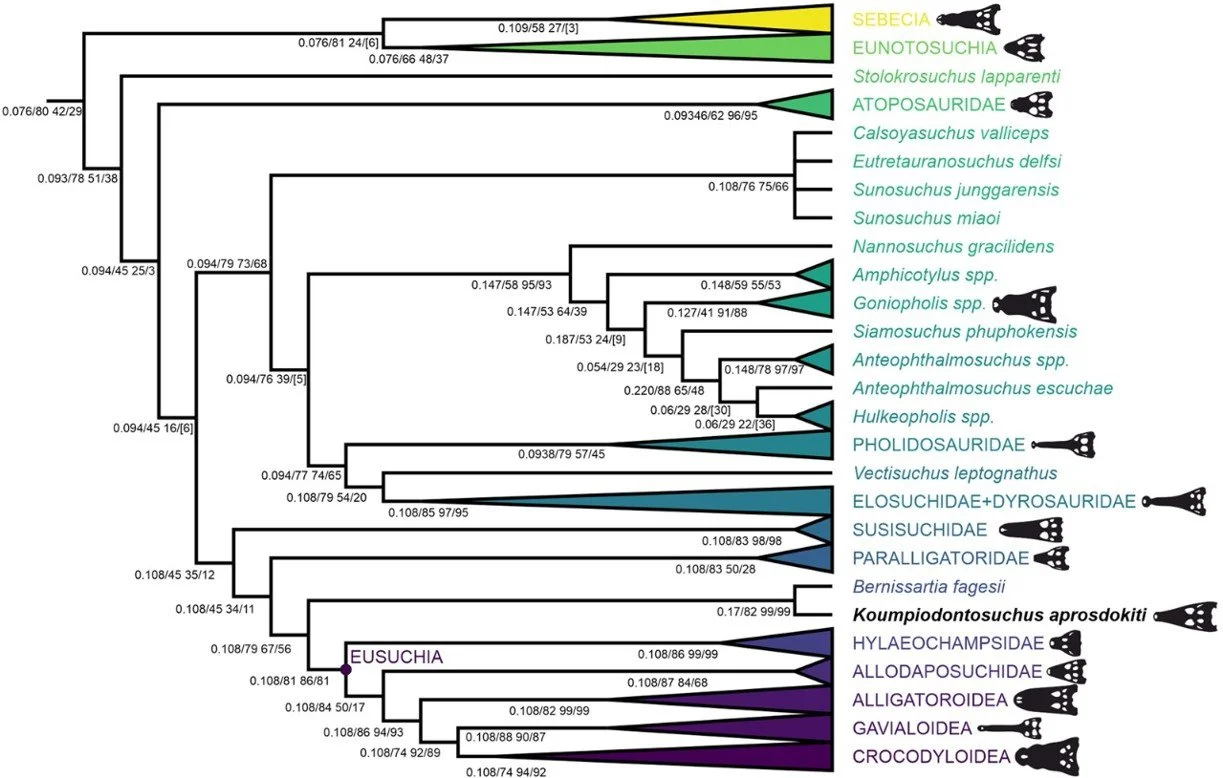
Caption: a phylogeny using extended implied weights, and with a more resolved topology. The goniopholidid + tethysuchian clade is again present. Images: Barker et al. (2026).
I haven’t used this article as an opportunity to discuss Wealden crocodyliforms in general, but it’s worth emphasising that Koumpi is but one of several, quite a few of which are also represented by excellent skull information (Salisbury & Naish 2011, Ristevski et al. 2018). Work similar to that done by our team on Koumpi could, one day, be done on those too and there are – as ever – more announcements on these animals yet to come…
Huge thanks and congrats to everyone involved in the Koumpi team, to everyone who assisted with this project at the University of Southampton and Dinosaur Isle Museum (where the fossil is housed), and to our phenomenally helpful and enthusiastic reviewers. The paper is OPEN ACCESS so please do check it out.

Caption: a montage of crocodylomorph-themed images, showing things that were published at Tet Zoo ver 2 and 3. There were articles there on thalattosuchians, recently extinct and extant crocodylians, and much more. Images: Tony Pyrakowski; Darren Naish.
Tet Zoo’s lament. A substantial amount of crocodylomorph material exists in the Tet Zoo archives but most of it is now only findable via the wayback machine and I haven’t yet added it to the WIP list, all of which means that I have to start from scratch and that virtually nothing is present here at ver 4, gah. Anyway, for previous articles at ver 4 on crocodylomorphs and Wealden animals in general, see…
Theropod Dinosaurs of the English Wealden, Some Questions (Part 1), March 2020
Dr Angela Milner and the Discovery of Baryonyx, August 2021
Two New Spinosaurid Dinosaurs from the English Cretaceous, August 2021
A Giant Spinosaurid Dinosaur from the Cretaceous of the Isle of Wight, June 2022
A brain for Baryonyx: using CT-scanning to examine British spinosaurid brains, February 2023
Discovering 'Hidden' Diversity Within Wealden Spinosaurid Dinosaurs, May 2023
A New Theropod Dinosaur Assemblage from the Older Part of the English Wealden, December 2024
Crocodiles Attack Elephants Then, Now, and Still, August 2025
Refs - -
Brochu, A. C., Wagner, J. R., Jouve, S., Sumrall, C. D. & Densmore, L. D. 2009. A correction corrected: consensus over the meaning of Crocodylia and why it matters. Systematic Biology 58, 537-543.
Martin, J. E., Smith, T., Salaviale, C., Adrien, J. & Delfino, M. 2020. Virtual reconstruction of the skull of Bernissartia fagesii and current understanding of the neosuchian–eusuchian transition. Journal of Systematic Palaeontology 18, 1079-1101.
Naish, D. & Sweetman, S. C. 2026. A second specimen of the enigmatic Wealden reptile Yaverlandia. Proceedings of the Geologists’ Association https://doi.org/10.1016/j.pgeola.2026.101179
Ristevski, J., Young, M. T., Brandalise de Andrade, M. & Hastings, A. K. 2018. A new species of Anteophthalmosuchus (Crocodylomorpha, Goniopholididae) from the Lower Cretaceous of the Isle of Wight, United Kingdom, and a review of the genus. Cretaceous Research 84, 340-383.
Salisbury, S. W. & Naish, D. 2011. Crocodilians. In Batten, D. J. (ed.) English Wealden Fossils. The Palaeontological Association (London), pp. 305-369.



