The rumours are true! At last, we have data on baryonychine brains…

Caption: the Isle of Wight baryonychine spinosaurid Ceratosuchops interacts with another animal, with the position and anatomy of its brain (and associated structures) shown in the inset. Image: Anthony Hutchings.
Long, long ago – in 2021 – myself and a team of colleagues (mostly associated with the University of Southampton) published our initial descriptive volley on the new British spinosaurid dinosaurs Ceratosuchops inferodios and Riparovenator milnerae (Barker et al. 2021). The study was led by Chris Barker, and analysis of the two animals forms the bulk of his PhD work.
Ceratosuchops and Riparovenator are both from the Lower Cretaceous Wessex Formation (part of the famous, and famously complex, Wealden Supergroup) of the Isle of Wight, and both lived during the Barremian age of the Early Cretaceous, round about 127 million years ago. Neither is complete nor even close to it, but both include well-preserved sections of skeletal anatomy. The braincases of both are three-dimensional and well preserved, as is also the case for the related Baryonyx walkeri from the Upper Weald Clay Formation of the English mainland (Charig & Milner 1986, 1997). All three are united within the spinosaurid group Baryonychinae.
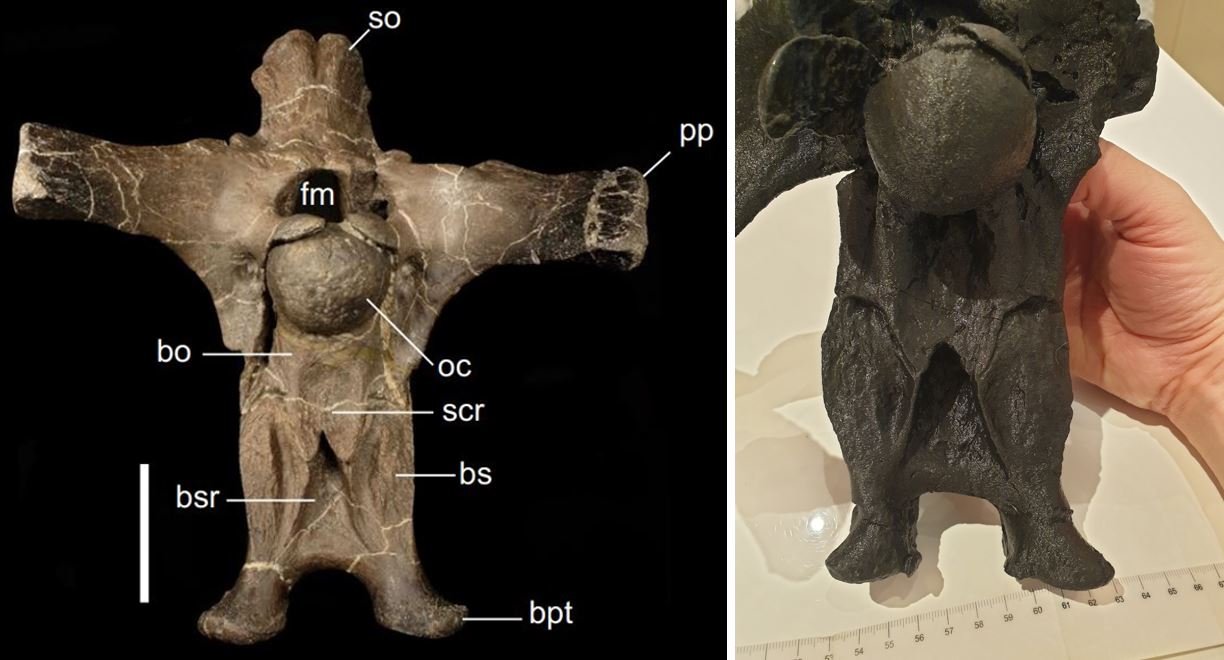
Caption: it should be obvious how well-preserved the remains of Ceratosuchops and Riparovenator are. These photos both show posterior surfaces of the braincases: at left is the original of Ceratosuchops (scale bar = 50 mm); at right, a 3D print of Riparovenator. Images: Barker et al. (2021); Darren Naish.
I’ve mentioned a few times that additional work on the Isle of Wight baryonychines is due to appear this year and beyond, and today sees another of our publications on these dinosaurs: ‘Modified skulls but conservative brains? The palaeoneurology and endocranial anatomy of baryonychine dinosaurs (Theropoda: Spinosauridae)’; it’s published in Journal of Anatomy (Barker et al. 2023). Chris and his PhD supervisor Neil Gostling led this research, and the contributors in addition to myself are Jacob Trend, Lysanne Veerle Michels, Larry Witmer, Ryan Ridgley, Katy Rankin, Claire Clarkin and Philipp Schneider.
What work did we do, and what are the conclusions?
Let’s CT-scan some baryonychines. Anyone paying attention to the scientific study of extinct dinosaurs will be aware of the revolution that computed tomography (or CT) has brought to the field. Thanks to advances made in CT-scanning technology and in the availability of CT-scanners themselves, scientists interested in the biology, behaviour, function and evolution of extinct animals now regularly place fossils in CT-scanners and analyse the results (Witmer et al. 2008). We’re especially blessed at the University of Southampton since we have the giant walk-in machine at the μ-VIS X-Ray Imaging Centre.

Caption: working with giant CT-scanners is not for for the feint of heart. This photos isn’t from the baryonychine project, but from another dating to 2012. Image: Darren Naish.
A caveat worth pointing out is that fossils from certain stratigraphic units tend not to scan well: the composition and density of the sediment that often surrounds and infills the fossils ruins the scanner’s ability to pick out fossil bone. This concern is especially relevant to Wealden fossils, since some of the respective sediments are full of minerals that disrupt easy scanning efforts. Exactly this affects one of the two Isle of Wight spinosaurids: the Riparovenator specimen is chock full of radio-opaque material that essentially make it impenetrable for this sort of work.
But I’m pleased to report that this is not the case for all Wealden spinosaurid specimens. We scanned the Ceratosuchops holotype in Southampton and the Baryonyx holotype at the Ohio Health O'Bleness Hospital in Athens, Ohio, and the key novelty of our study is that we provide the first CT-scan data yet reported for baryonychines (Barker et al. 2023). Both yield good results, and we reasoned that the good data on brain anatomy that they revealed would prove informative with respect to baryonychine biology and lifestyle. As discussed below, these aren’t the first spinosaurids to be CT-scanned, but they are the first baryonychines.

Caption: image showing the brain of Baryonyx in-situ inside the braincase. The braincases of baryonychines are narrow and deep, with basipterygoid processes that descend way below the rest of the structure. The section of the brain we’re seeing here (it doesn’t include the olfactory bulbs and tract) is about 12.8 cm long. Image: WitmerLab and Chris Barker.
Baryonychines vs other theropods. What do the scans mean with respect to how baryonychine brains compare to those of other theropods? We do at least have brain data for other, related theropods, including one other spinosaurid, as well as members of the more distantly related Allosauroidea. Here I should remind you that megalosaurids and spinosaurids are close kin within the theropod clade Tetanurae, both forming the clade Megalosauroidea. Views differ as to whether allosauroids are more closely related to birds than megalosauroids are, or whether there’s a megalosauroid + allosauroid clade.

Caption: the first of two competing theropod cladograms, highly simplified. This tree shows the more ‘traditional’ topology (at least, in these post-Gauthier 1986 times) in which allosauroids are closer to coelurosaurs than to megalosauroids (e.g., Holtz et al. 2004, Carrano et al. 2012). Image: Darren Naish.

Caption: a tree found in some studies (e.g., Rauhut 2003, Naish & Cau 2022), where megalosauroids and allosauroids form a clade. Image: Darren Naish.
In overall structure and proportion, baryonychine brains conform to what we’d expect for theropods of this sort. The forebrain and hindbrain are in line with one another in the horizontal plane, there’s an elongate, narrow olfactory tract that projects forwards to the olfactory bulbs, the optic lobes are difficult to differentiate from the rest of the forebrain, the floccular lobes are present as projecting tabs surrounded by the semicircular canals of the inner ear, and the cranial nerves are in the expected position. The braincases of both animals are pneumatic, but not to the degree present in coelurosaurs (Barker et al. 2023).
The brains of Ceratosuchops and Baryonyx differ in several respects. Ceratosuchops has a midline groove along its upper surface, the two differ in terms of the position and extent of the peaked section at the top of the cerebrum (it’s less prominent and located a bit further back in Ceratosuchops), the Ceratosuchops forebrain is inclined downwards (whereas that of Baryonyx is more horizontal), the Baryonyx inner ear is proportionally larger than that of Ceratosuchops, and both differ in the shape of the lateral semicircular canal (Barker et al. 2023).

Caption: a baryonychine brain CT-scan composite, featuring a brain scan of Baryonyx walkeri at left, and Ceratosuchops inferodios at right. Image: Barker et al. (2023).
Why so conservative? Overall, these brains aren’t tremendously different from those of other large tetanuran theropods. If anything, baryonychine brains appear anatomically conservative, certainly so relative to the bony anatomy of their skulls. That’s a surprising result, since the modification and specialisation of the spinosaurid skull – the rostrum in particular – might lead you to predict a modified brain too.
There are several ways to interpret these results. One possibility is that brain evolution was ‘lagging behind’ the evolution of the bony parts of the skull, either because brains and the nervous system are slow to evolve or conservative overall, or because the ancestral morphology was ‘good enough’ and that further modification wasn’t needed. Another possibility is that the megalosaurid-like theropods ancestral to spinosaurids already possessed the sensory and somatosensory features required for a successful baryonychine lifestyle. In other words, megalosauroids were pre-adapted for aquatic predation and an amphibious lifestyle, and ‘all’ they needed to do to become adept at it was modify the snout, jaws and teeth (Barker et al. 2023).
Other possible explanations exist though. A third possibility is that changes had happened, but concern details of brain structure that aren’t knowable via CT-scan data. And a fourth possibility is that baryonychines just didn’t need specialised brains relative to other theropods because all this speculation about them being heron-like waders is overblown. Maybe they were ecological generalists, predating on all manner of aquatic and terrestrial prey, and not much different in foraging behaviour from megalosaurids and the like. That proposal does a poor job of explaining why their skulls are otherwise so modified, however, so I have a hard time buying it.

Caption: a reconstructed, mounted skeleton of the spinosaurine Irritator, as previously displayed at the National Museum of Rio de Janeiro, and posed with an anhangeurid pterosaur as a prey item. This photo was taken in 2013. Image: Darren Naish.
I mentioned earlier than we have brain data from one other spinosaurid. In 2020, Marco Schade and colleagues reported CT-scan data for the brain of Irritator, a South American spinosaurine spinosaurid (Schade et al. 2020). One aspect in which it differs from the baryonychines (and other related theropods too) is that the floccular lobe is larger. Because the floccular lobe is relevant to gaze stabilisation via coordination of head, eye and neck movement, the large lobes of Irritator could be a specialisation for aquatic predation (Schade et al. 2020). Conversely, the more ‘typical’ floccular lobe of baryonychines could mean that they were less specialised, or non-specialised, for aquatic predation (though, another caveat: there’s some controversy as to how reliable a guide floccular lobe is for ecology and behaviour).

Caption: the hindbrain region of (a) Baryonyx and (b) Ceratosuchops, showing the arrangement of cranial nerves (in yellow), the semicircular canals (in pink), and the associated segment of cerebrum (in blue). The right side’s floccular lobe – labelled fl – is tab-shaped and projects into the semicircular canals. Image: Barker et al. (2023).
Some specifics, and some implications. As should be obvious from the paper, we did just about everything we could from the data we gleaned. We estimated encephalisation quotient (EQ)*, hearing range, olfactory acuity and possible head pose.
* Because it’s been shown that vertebrates as a whole don’t fit on the same brain : body size coefficient, experts today use different ones for the different vertebrate group. That for non-bird reptiles is termed REQ (Hurlburt et al. 2013).
By comparing estimated brain size to body size, we can work out the REQ of our sampled baryonychines, and it’s between 1.2 and 1.6 (Barker et al. 2023). That’s in the same approximate range as that of theropods like Carnotaurus and Allosaurus and could mean that these animals were similar in cognitive abilities and behavioural complexity (and here I’ll avoid any further discussion of how reliable a guide REQ is to ‘intelligence’, for reasons).

Caption: these days, there’s often a fairly rapid turnover time between the publication of a new dinosaur taxon and the appearance of good, commercially available figures of said taxon. New for 2023 is this Ceratosuchops figure from CollectA which I’m very pleased to own in my collection. Images: Darren Naish.
Moving on… based on cochlear duct size relative to basicranial length, it looks like Ceratosuchops and Baryonyx had mean hearing frequencies in the 1400-1600 Hz range (Barker et al. 2023). That’s very approximately in the crocodylian range but also – for those who might think that this is significant – in about the same range as a great many birds. On olfaction, there’s been some suggestion that spinosaurids had reduced olfactory bulbs and thus a reduced sense of smell, but this doesn’t seem to be the case in baryonychines at all. Their olfactory bulbs are ‘normal’ for theropods of their size, and their sense of smell was likely pretty good, and again about similar to that of abelisaurids and Allosaurus (Barker et al. 2023). Finally, both the nature of the condyle at the back of the skull and orientation of the semicircular canals imply that baryonychines were ‘standard’ as theropods go (Barker et al. 2023). There’s no indication that they maintained the skull at a dipped-down angle or anything like that, as has been inferred for Irritator (Schade et al. 2020).
All in all, our data “indicates that baryonychines did not deviate substantially from the cognitive, auditory or olfactory capabilities of ‘typical’ non-coelurosaurian theropods” (Barker et al. 2023, p. 17). Interesting, and weird.
Caption: life reconstruction of Ceratosuchops (at left) and Riparovenator, shown sharing a flooded depression on the Wessex Formation floodplain. Image: Anthony Hutchings.
And right now that sums up just about everything that’s worth saying at this point. Just successfully obtaining CT data on brain anatomy – palaeoneurological data – from baryonychine spinosaurids is a big deal, and using it to infer sensory abilities and so on is also a worthy endeavour. Finding that baryonychines appear surprisingly samey relative to non-spinosaurid theropods is a curious result, but as knowledge improves we’ll understand better just what this means. I want to thank Chris, Neil, Jacob and my other coauthors for their co-operation on such an interesting study, and I’m pleased that we’ve succeeded in contributing once again to our growing knowledge of Wealden spinosaurids.
On that note -- more Wealden spinosaurid news is coming!
For previous TetZoo articles on spinosaurids, British theropods and associated issues (some links here are to wayback machine versions due to destruction or paywalling of everything at versions 2 and 3), see…
Of Becklespinax and Valdoraptor, October 2007
The world’s most amazing sauropod, November 2007
Oh no, not another new Wealden theropod!, June 2009
Concavenator: an incredible allosauroid with a weird sail (or hump)... and proto-feathers?, September 2010
The Wealden Bible: English Wealden Fossils, 2011, November 2011
Ostrich dinosaurs invade Europe! Or do they?, June 2014 (every archived version of this article lacks the original illustrations, sorry)
Theropod Dinosaurs of the English Wealden, Some Questions (Part 1), March 2020
Introducing ‘Unexpected Isle of Wight Air-Filled Hunter’, a New English Theropod Dinosaur, September 2020
Dr Angela Milner and the Discovery of Baryonyx, August 2021
Two New Spinosaurid Dinosaurs from the English Cretaceous, September 2021
A Giant Spinosaurid Dinosaur from the Cretaceous of the Isle of Wight, June 2022
Refs - -
Carrano, M. T., Benson, R. B. J. & Sampson, S. D. 2012. The phylogeny of Tetanurae (Dinosauria: Theropoda), Journal of Systematic Palaeontology 10, 211-300.
Charig, A. J. & Milner, A. C. 1986. Baryonyx, a remarkable new theropod dinosaur. Nature 324, 359-361.
Charig, A. J. & Milner, A. C. 1997. Baryonyx walkeri, a fish-eating dinosaur from the Wealden of Surrey. Bulletin of the Natural History Museum 53, 11-70.
Holtz, T. R., Molnar, R. E. & Currie, P. J. 2004. Basal Tetanurae. In Weishampel, D. B., Dodson, P. & Osmólska, H. (eds) The Dinosauria, Second Edition. University of California Press (Berkeley), pp. 71-110.
Hurlburt, G. R., Ridgely, R. C. & Witmer, L. M. 2013. Relative size of brain and cerebrum in tyrannosaurid dinosaurs: an analysis using brain endocast quantitative relationships in extant alligators. In Parrish, J. M., Molnar, R. E., Currie, P. J. & Koppelhus, E. B. (eds.) Tyrannosaurid Paleobiology. Bloomington, Indiana University Press, pp. 134-154.
Rauhut, O. W. M. 2003. The interrelationships and evolution of basal theropod dinosaurs. Special Papers in Palaeontology 69, 1-213.
Witmer, L. M., Ridgely, R. C., Dufeau, D. L. & Semones, M. C. 2008. Using CT to peer into the past: 3D visualization of the brain and ear regions of birds, crocodiles, and nonavian dinosaurs. In Endo, H. & Frey, R. (eds) Anatomical Imaging. Berlin, Springer, pp. 67-87.



