Inspired by the recent publication here of thoughts on the new Koumpiodontosuchus paper… and by other work, in prep… I felt it appropriate to rescue another article from the archives, specifically from ver 3. Here we go…
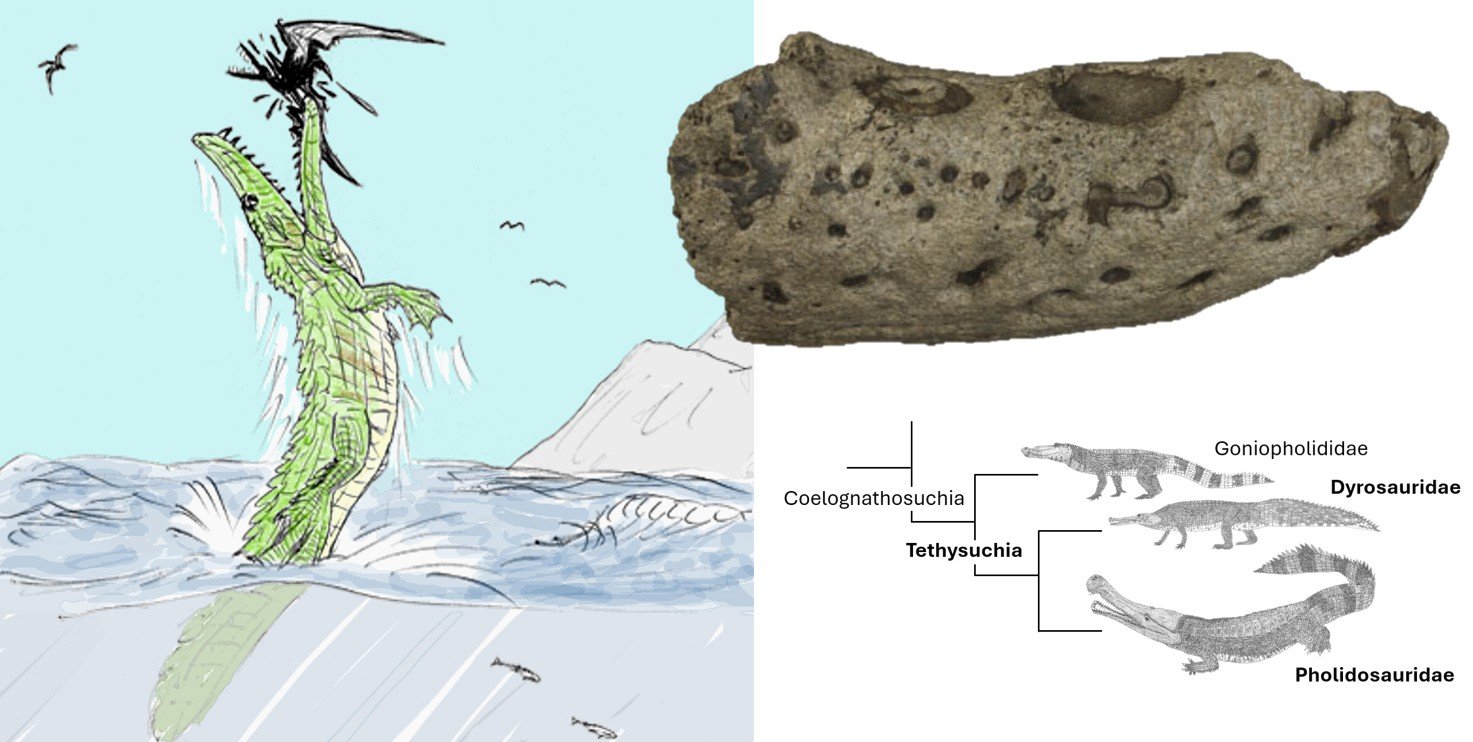
Caption: a Shanklin croc montage, showing the actual fossil at upper right. At left is a not entirely serious reconstruction by Vladimir Dinets (colourised by Darren Naish).
The article concerned was published in 2014 (it’s here at wayback machine) and revolves around the publication of a technical paper co-authored by myself and colleagues, led by marine crocodylomorph guru Mark Young (Young et al. 2014).
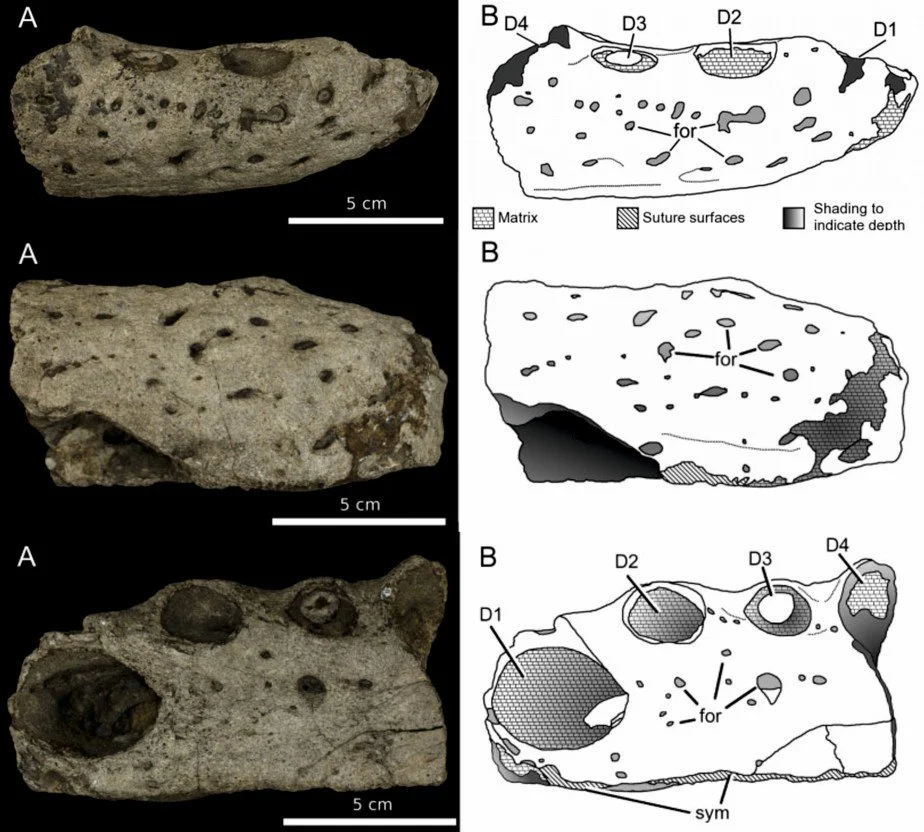
Caption: the Shanklin croc specimen (NHMUK PV OR36173) in (at top) right lateral, (at middle) ventral, and (at bottom) dorsal view. Images from Young et al. (2014). It's not the prettiest fossil in the world, but it's still interesting.
This paper describes another of those frustrating fossils that combines anatomical novelty and possible phylogenetic significance with a most unsatisfactory degree of completeness. The fossil concerned is an ugly chunk of bone (going by the catchy accession number NHMUK PV OR36173), about 13 cm long, that represents the anterior part of the incomplete right dentary bone of a reasonably large, long-snouted Cretaceous crocodylomorph. It comes from Shanklin on the south-east coast of the Isle of Wight, England. While my innovative co-author Lorna Steel proposed the nickname Shanklin Shocker for the creature, I’ll simply refer to it from hereon as the ‘Shanklin croc’.

Caption: Richard Lydekker (1849-1915): biogeographer, anthropologist, zoologist, palaeontologist, here photographed in or around 1900. Image: (c) but allowed under fair use. Original here.
History and provenance. The specimen isn’t completely new to the literature. We know that it was purchased by the then British Museum (Natural History) (now the Natural History Museum, or NHM) in 1861 from a Mr Simmons. Palaeontologist, zoologist and famed ‘lightning cataloguer’ Richard Lydekker then wrote about it in 1889 but (mis)identified it as the premaxillary bone of the plesiosaur Polyptychodon. Fast forward about a century, and Leslie Noé – known to marine reptile workers for his (still predominantly unpublished) PhD work on pliosaurids – left a note in the NHM collections saying that it isn’t a plesiosaur at all, but more likely a crocodylomorph of some sort.
It’s this that brought it to the attention of croc-worker Lorna Steel; Lorna has been working for some years with Mark Young on new (and/or previously overlooked or under-appreciated) crocodylomorphs. Mark and Lorna then assembled a team of people who work on relevant crocodylomorph groups. I became involved because we needed to determine whether the specimen was anything to do with the goniopholidids, a group of long-snouted Jurassic and Cretaceous crocodyliform crocodylomorphs well represented in the fossil record of the Isle of Wight. Regular readers will perhaps recall that Steve Salisbury and I reviewed the Wealden crocodyliforms back in 2011 and therein named or renamed several new species (Salisbury & Naish 2011). This isn’t the first time crocodylomorph remains have been identified as those of a plesiosaur, by the way (e.g., Buchy 2008).
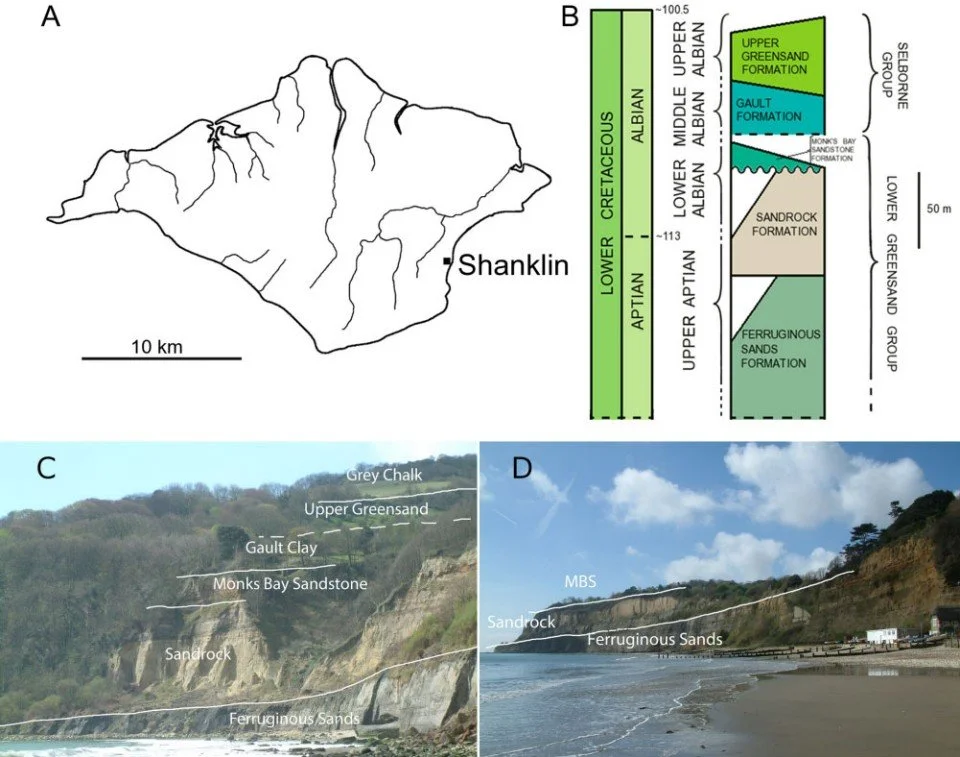
Caption: locational and stratigraphic data on the Shanklin croc. It comes from the south-east coast of the Isle of Wight (C shows Luccombe Chine, D shows Knock Cliff), though exactly which Cretaceous stratum it came from is uncertain. Knock Cliff has also yielded a fragmentary theropod that I’ve worked on, namely the holotype of Vectaerovenator inopinatus Barker et al., 2020. This figure is from the paper (Young et al. 2014).
The precise provenance of the Shanklin croc is somewhat uncertain and there are several different sedimentary strata it could have come from. Based on the fossil’s colour, it seems most likely that it came from the ‘Malm rock’ of the Upper Greensand Formation, a geological unit deposited during the late Albian during the Early Cretaceous. A few other units that crop out in the same area and are Aptian or Albian in age are also possible sources for the specimen, meaning that we’ve given its age as ‘?Aptian-Albian’ (Young et al. 2014). Upper Albian seems most likely.
Incidentally, if you’re confused as to why I’m seemingly inconsistent in my use of ‘lower’/‘upper’ and ‘early’/‘late’, it’s because ‘lower’ and ‘upper’ refer to the position of rock layers while ‘early' and ‘late’ refer to geological time. So, the Shanklin croc lived during the late Albian of the Early Cretaceous, but its fossil was found in the upper Albian of the Lower Cretaceous.
What sort of crocodylomorph might the Shanklin croc be? Could it belong to any of the groups already reported from the Isle of Wight Cretaceous? These are atoposaurids, goniopholidids, bernisartiids, and hylaeochampsids. Short answer: no. Long answer: read the paper [contact any of the authors if you want a pdf]. Intermediate-length answer: as revealed by comparisons made between the Shanklin croc and the dentaries of those other groups, none possess the specific alveolar configuration or mandible shape that the Shanklin croc does and all possess anatomical peculiarities not present in the Shanklin croc (Young et al. 2014). We could also exclude several other Cretaceous crocodylomorph groups, including gavialoids, teleosaurids, metriorhynchids and the several lineages often grouped together in Pholidosauridae (Young et al. 2014). So... what is it?
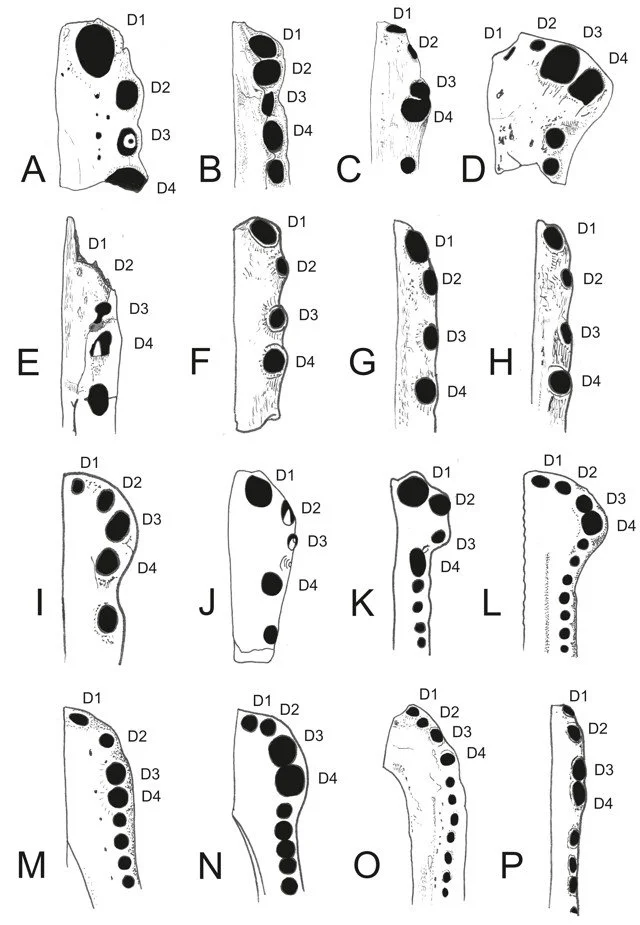
Caption: the Shanklin croc specimen (A) compared with the anterior dentaries of assorted other crocodylomorphs, including thalattosuchians, dyrosaurids, goniopholidids, hylaeochampsids and gavialoids. Diagram from Young et al. (2014).
Based on the presence of enlarged anterior alveoli, the shape of the bone’s outer margin, the large and widely spaced foramina on the lateral and ventral surfaces of the bone and other features, it seems most similar to species included within Dyrosauridae, a group of long-snouted crocodyliforms of the Cretaceous and Paleogene. However, it remains unusual enough compared to undoubted dyrosaurids that we have some reservations about referring it to Dyrosauridae outright. Our conservative conclusion is that the Shanklin croc should be identified as Tethysuchia incertae sedis, Tethysuchia being the crocodyliform clade that includes dyrosaurids, pholidosaurids and allied taxa.
Meet the tethysuchians. Tethysuchians haven’t been much (if at all) discussed on Tet Zoo before, so this is a good time to say some things about them. Experts have long recognised several Jurassic, Cretaceous and Paleogene crocodylomorph lineages that all look roughly alike in being long-jawed, strongly aquatic predators that have a substantially ‘archaic’ osteoderm configuration compared to living crocodylomorphs (the crocodylians).
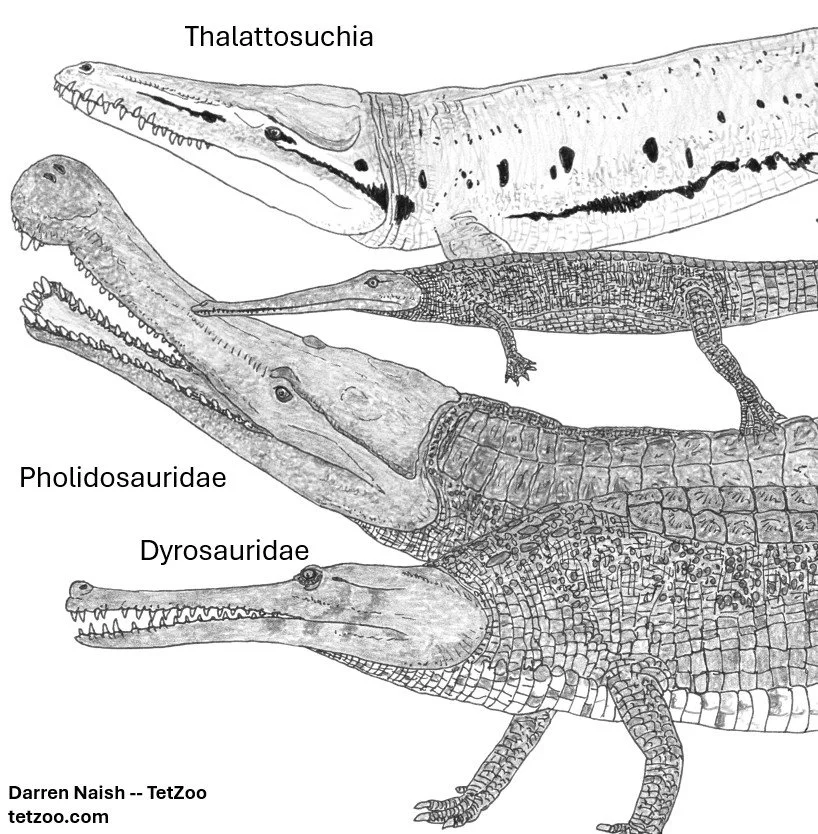
Caption: the three crocodylomorph groups originally assembled in the 'longirostrine clade' by Clark (in Benton & Clark 1988). Thalattosuchia includes the two main groups Teleosauroidea and Metriorhynchoidea. Images: Darren Naish.
At the core of the assemblage are the dyrosaurids of the latest Cretaceous and Paleogene and the pholidosaurids of the Early and Late Cretaceous. Benton & Clark (1988) and Clark (1994) grouped these together with the sea-going thalattosuchians into a ‘longirostrine clade’ (they didn’t give it an official name). Goniopholidids have also been allied with the ‘longirostrine clade’ in some studies (Andrade et al. 2012). Brand-new at the time of writing is Jouve et al.’s (2026) proposal that the name Coelognathosuchia be used for the goniopholidid + tethysuchian clade, and Stenorhynchosuchia for the thalattosuchian + tethysuchian clade. These names are mutually exclusive: Coelognathosuchia can only be used when goniopholidids are close to tethysuchians (it cannot include thalattosuchians), and Stenorhynchosuchia can only be used when thalattosuchians are close to tethysuchians (it cannot include goniopholidids) (Jouve et al. 2026).
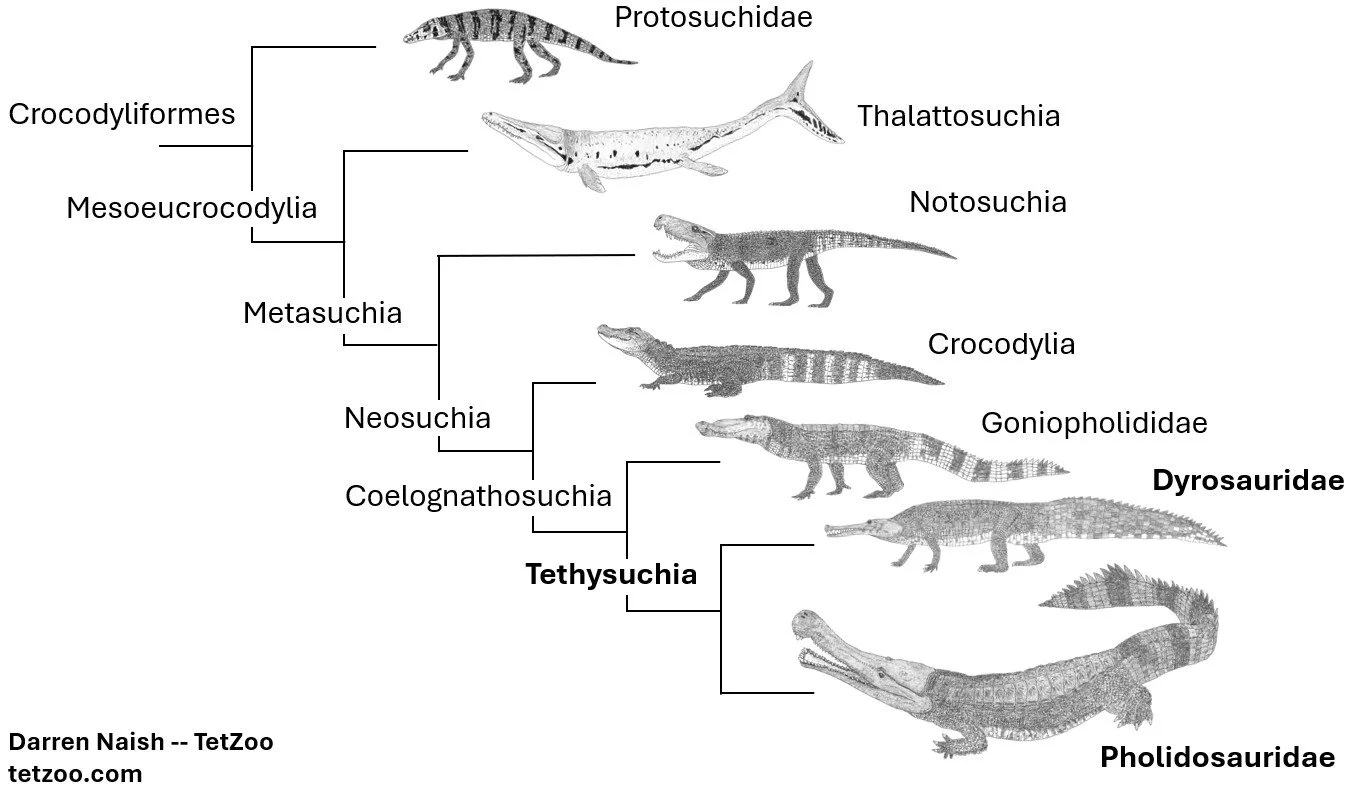
Caption: a highly simplified version of a phylogenetic hypothesis supported in several studies, where thalattosuchians are well away from crown-crocs (Crocodylia) and are not close to tethysuchians. This tree also shows goniopholidids close to Tethysuchians, in which case we could use the name Coelognathosuchia for the relevant clade. That name was first published by Martin et al. (2014) and recently given a phylogenetic definition by Jouve et al. (2026). Image by Darren Naish.
On that note, do all of these longirostrine crocodylomorphs really go together? Well, this is controversial. If thalattosuchians really are close relatives of dyrosaurids and pholidosaurids, this makes thalattosuchians ‘advanced’ crocodyliforms, not all that far in phylogenetic distance from crown-crocs. But other studies don’t support a close relationship between thalattosuchians, goniopholidids and the other longirostrine groups, instead finding thalattosuchians far, far away from the crown (Pol & Gasparini 2009, Sereno & Larsson 2009, Young et al. 2012, Montefeltro et al. 2013), and perhaps not even in Neosuchia at all. This story was summarised a while back at Tet Zoo [UPDATE: in another article now only findable at wayback machine]. Most character evidence puts goniopholidids closer to crown-crocs than are dyrosaurids and pholidosaurids (e.g., Salisbury et al. 2006, Montefeltro et al. 2013), but not all studies find this. Anyway, the longirostrine ‘core’ consists only of dyrosaurids, pholidosaurids and close kin.
Buffetaut (1982) was first to make the case for Tethysuchia and coined that name. Its use is widespread in crocodylomorph studies today. Pholidosaurids have sometimes been found to represent a paraphyletic series of outgroups to Dyrosauridae but more recent work has supported their monophyly.
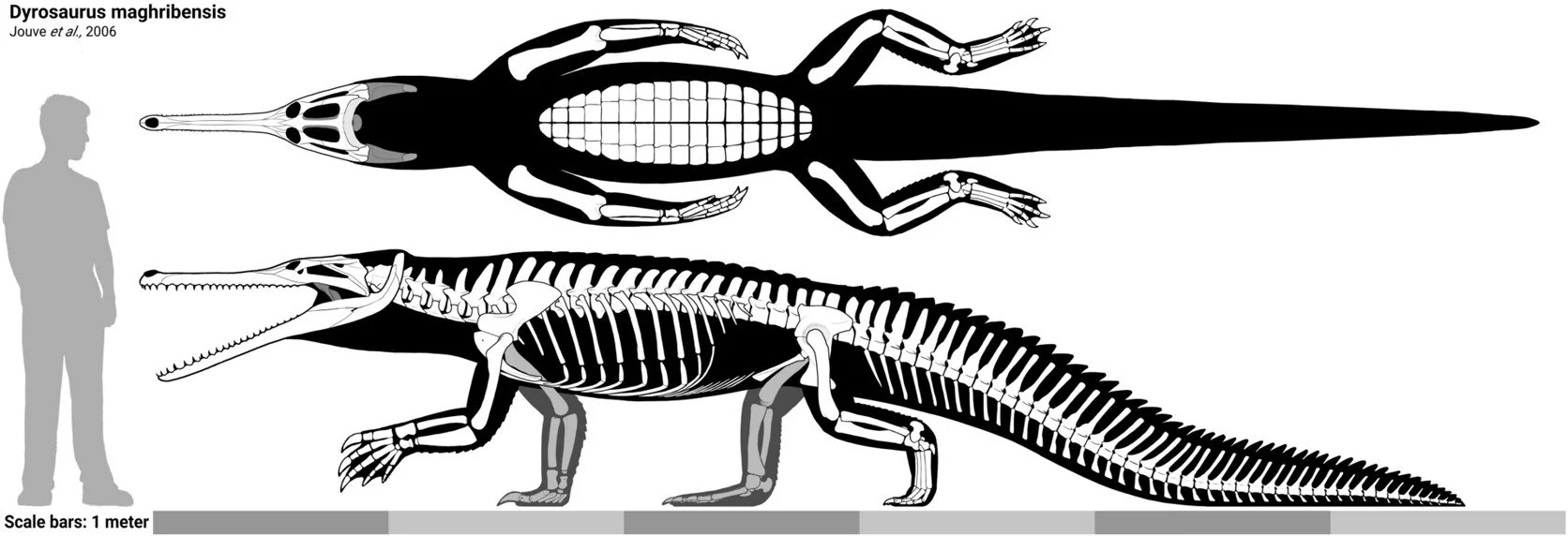
Caption: skeletal reconstruction of Dyrosaurus to scale with a person. Note the long supratemporal fenestrae and tall neural spines in the shoulder region. The very long, strongly curved retroarticular process (the curving structure at the posterior end of the lower jaw) is interesting too. Image: LiterallyMiguel, CC BY 4.0 (original here).
Thoughts on dyrosaurids. Dyrosaurids haven’t really had their monophyly doubted and are now a large and complex group. They’re predominantly associated with Africa (and likely originated there) but species also inhabited southern Asia and both North and South America. They were generally slender-snouted animals that grabbed fish but some (like giant Phosphatosaurus from the Eocene of Tunisia) have robust jaws and blunt teeth and look capable of handling robust prey items. Others (like Cerrejonisuchus from the Paleocene of Colombia) are especially robust-snouted and yet others (like Anthracosuchus, also from the Paleocene of Colombia) are comically short-snouted.
Dyrosaurid skulls are reasonably easy to recognise. The eye sockets are dorsally placed and the supratemporal openings are about twice as long as they are wide. Dyrosaurids represented by good postcranial remains reveal unusual proportions relative to those of crocodylians. The forelimbs are long, slender and sometimes longer than the hindlimbs, tall neural spines on the dorsal vertebrae must have created a deep chest and somewhat hump-backed appearance and the tall is especially deep and narrow.
Schwarz-Wings et al. (2009) reconstructed dyrosaurid musculature and reported evidence for unusually large and strong limb, body and tail musculature. They suggested that dyrosaurids below a certain body size might have been especially strong walkers, that their ability to generate thrust by the tail exceeded that of crocodylians, and that their powerful body and tail allowed them to better move and forage in strong currents. These features could conceivably have been specialisations for life in tidal, coastal habitats. Not all dyrosaurids are alike though: the relatively small, short-snouted Cerrejonisuchus has features suggestive of a semi-terrestrial way of life (Scavezzoni & Fischer 2021).
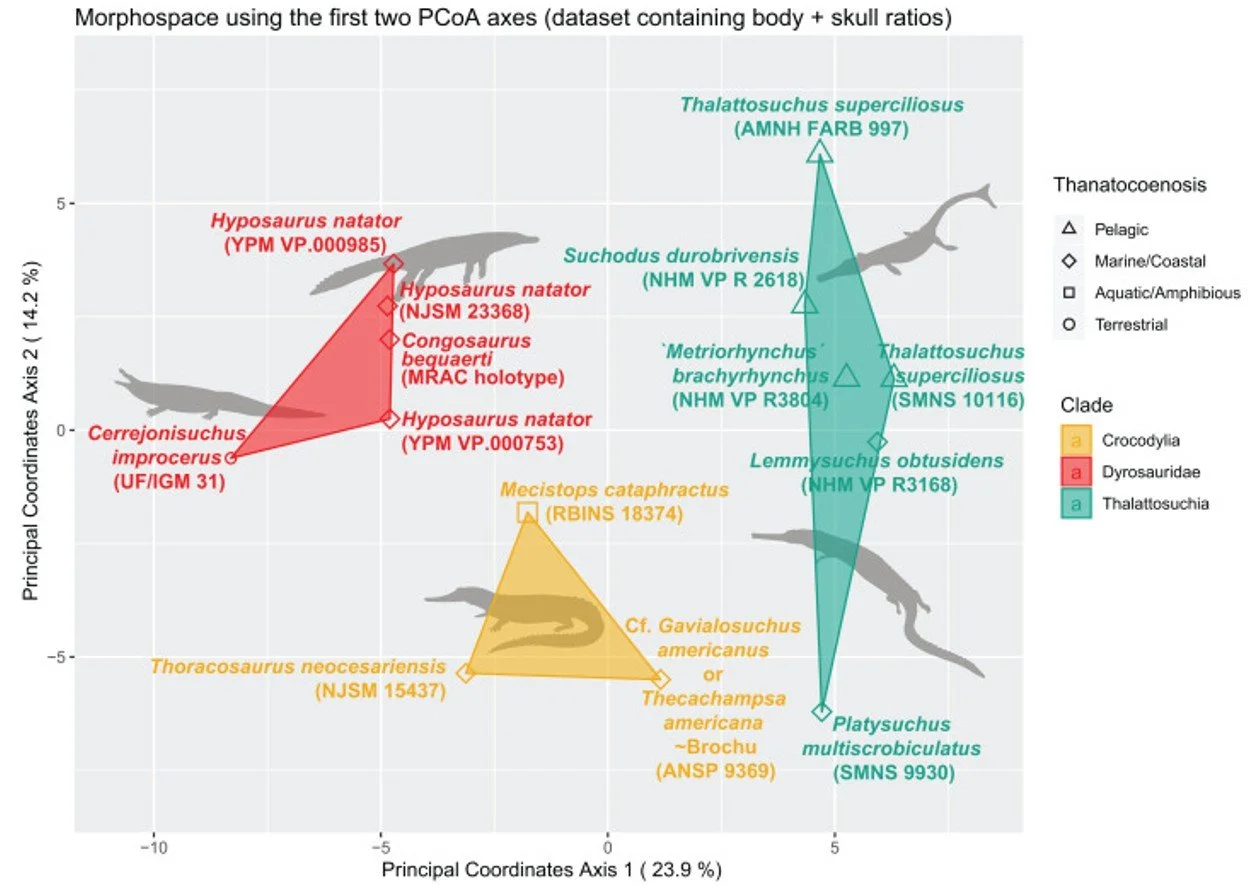
Caption: as is the case for so many extinct crocodylomorph groups, dyrosaurids are actually not that similar to crocodylians when we really pay attention to their proportions and various traits. This graph, from Scavezzoni & Fischer (2021), shows how dyrosaurids (red) differ notably from crocodylians (yellow) in anatomical proportions, as do thalattosuchians (green). Image: Scavezzoni & Fischer (2021), CC BY 4.0 (original here).
What, then, to make of the Shanklin croc? Our identification of the Shanklin croc as some sort of dyrosaurid-like tethysuchian makes it especially interesting, because it’s geologically older than expected based on what we know about tethysuchian history. Dyrosaurids are well known from the Maastrichtian onwards but pre-Maastrichtian specimens – some have been reported from the Cenomanian and Campanian (Buffetaut et al. 1990, Churcher & Russell 1992, Churcher 1995) – are fragmentary. Furthermore, the skull fragments concerned resemble those of some pholidosaurids and thus might not be from dyrosaurids after all; Buffetaut et al. (1990) did report some vertebrae that look very dyrosaurid-like, however. We conclude for now that there are no definite records of dyrosaurids in the ‘middle’ Cretaceous, the implication from the fragmentary remains reported so far – the Shanklin croc included – being that several ‘proto-dyrosaurid’-type taxa might have been in existence from the Aptian/Albian onwards, but that better remains are needed before we can say anything more.
As for which specific tethysuchian taxon the Shanklin croc represents, it’s something new: a new species and genus that needs a name. You might argue that we should have named it in the paper (part of me thinks that we should have). However, people are generally discouraged from attaching names to fragmentary specimens such as this. As ever, we hope that newer and substantially better material will turn up in time and that better remains from this part of the Cretaceous will help us pin down the origins and biogeographical history of one of the most important of marine crocodylomorph radiations.
Caption: stratigraphic ranges of relevant Lower Cretaceous crocodylomorph lineages, plotted against time and compared with the possible age range of the Shanklin croc. Dotted lines represent parts of lineages for which fossils are unreported; arrows show that the lineage concerned persisted beyond the end of the Turonian. Diagram from Young et al. (2014).
Your usual lament. Crocodylomorphs of several sorts have been covered here on a reasonable number of occasions but most of this material is now lost due to the death of ScienceBlogs and Scientific American blogs. Consequently, only a small number of articles are easily findable today. I aim to recover and republish my older articles here, in time. Sigh. Anyway…
Crocodiles Attack Elephants Then, Now, and Still, August 2025
Koumpi the Wealden Croc: Tons of New Data and Phylogeny Resolved, April 2026
Refs - -
Andrade, M. B., Edmonds, R., Benton, M. J. & Schouten, R. 2012. A new Berriasian species of Goniopholis (Mesoeucrocodylia, Neosuchia) from England, and a review of the genus. Zoological Journal of the Linnean Society 163, S66–S108.
Benton, M. J. & Clark, J. M. 1988. Archosaur phylogeny and the relationships of the Crocodylia. In Benton, M. J. (eds) The Phylogeny and Classification of the Tetrapods, Volume 1: Amphibians, Reptiles, Birds. Clarendon Press (Oxford), pp. 295-338.
Buffetaut, E. 1982. Radiation évolutive, paléoécologie et biogéographie des crocodiliens mésosuchiens. Mémoires de la Société Géologique de France 60, 1-85.
Buffetaut, E., Bussert, R. & Brinkmann, W. 1990. A new nonmarine vertebrate fauna in the Upper Cretaceous of northern Sudan. Berliner Geowissenschaftliche Abhandlungen A 120, 183-202.
Churcher, C. S. 1995. Giant Cretaceous lungfish Neoceratodus tuberculatus from a deltaic environment in the Quseir (=Baris) Formation of Kharga Oasis, Western Desert of Egypt. Journal of Vertebrate Paleontology 15, 845-849.
Churcher, C. S. & Russell, D. A. 1992. Terrestrial vertebrates from Campanian strata in Wadi el-Gedid (Kharga and Dakhleh Oases), Western Desert of Egypt. Journal of Vertebrate Paleontology 12 (Supplement), 23A.
Clark, J. M. 1994. Patterns of evolution in Mesozoic Crocodyliformes. In Fraser, N. C. & Sues, H.-D. (eds) In the Shadow of the Dinosaurs – Early Mesozoic Tetrapods. Cambridge University Press (Cambridge, NY, Melbourne), pp. 84-97.
Jouve, S., Young, M. T., Hastings, A. K. & Salih, K. 2026. The systematics and nomenclature of Tethysuchia (Archosauria: Crocodylomorpha) under the International Code of Phylogenetic Nomenclature. Zoological Journal of the Linnean Society 206, zlag045.
Montefeltro, F. C., Larsson, H. C. E., de França, M. A. G. & Langer, M. C. 2013. A new neosuchian with Asian affinities from the Jurassic of northeastern Brazil. Naturwissenschaften 100, 835–841.
Pol, D. & Gasparini, Z. 2009. Skull anatomy of Dakosaurus andinensis (Thalattosuchia: Crocodylomorpha) and the phylogenetic position of Thalattosuchia. Journal of Systematic Palaeontology 7, 163-197.
Schwarz-Wings, D., Eberhard Frey, E. & Martin, T. 2009. Reconstruction of the bracing system of the trunk and tail in hyposaurine dyrosaurids (Crocodylomorpha; Mesoeucrocodylia). Journal of Vertebrate Paleontology 29, 453-472.
Young, M. T., Steel, L., Foffa, D., Price, T., Naish, D. & Tennant, J. P. 2014. Marine tethysuchian crocodyliform from the ?Aptian-Albian (Lower Cretaceous) of the Isle of Wight, UK. Biological Journal of the Linnean Society 113, 854-871.



