For some time now I’ve been interested in what the glyptodonts – a group of highly modified, large or gigantic armadillos – looked like when alive….

Caption: excellent Glyptodon museum display in Museum für Naturkunde, Berlin. Showing a ‘naked’ glyptodont alongside its carapace and tail armour is, at this point, a familiar trope. Image: Dellex, CC BY-SA 3.0 (original here).

Caption: a montage of glyptodont reconstructions, giving some idea of the diversity of views that exist on the life appearance of these animals. There’s considerable variation in snout shape and nostril position, in the external morphology of the osteoderm-supported region, and in how hairy or naked-skinned the animals are. Clockwise from upper left, these images are by J. Smit, Michael Long, Peter Schouten, and Margaret Lambert.
As discussed in those articles, glyptodonts are part of the armadillo group conventionally termed Chlamyphoridae (though… a new taxonomy is needed), and as such are cousins of euphractines (the hairy armadillos and Pichi), fairy armadillos and tolypeutines (the giant armadillo, three-banded armadillos and naked-tailed armadillos). Both molecular data and anatomy indicate that glyptodonts are the sister-group to the fairy armadillo + tolypeutine clade, in which case they’re phylogenetically bracketed by this lot on one side, and by euphractines on the other (Billet et al. 2011, Delsuc et al. 2016, Mitchell et al. 2016). Long-nosed armadillos (Dasypodidae) are distant cousins.
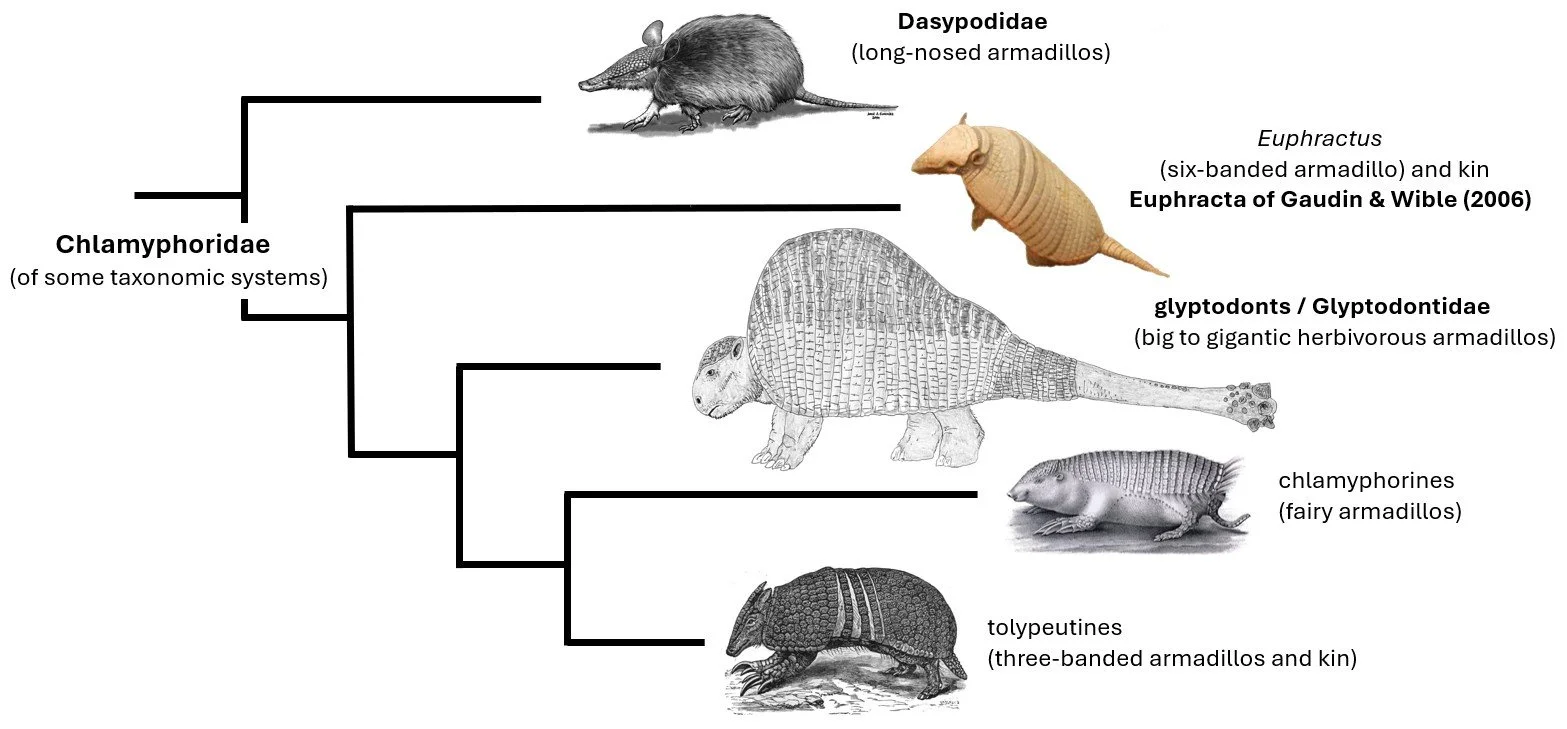
Caption: much simplified cladogram showing how glyptodonts are bracketed by living armadillos, namely euphractines on one side and fairy armadillos and tolypeutines on the other. Images: Dasypus by Jorge González from Castro et al. (2015); Euphractus and Doedicurus by Darren Naish; fairy armadillo by Hermann Burmeister, in public domain; three-banded armadillo in the public domain (original here).
Living armadillos are diverse in appearance. All possess horny scutes that grow over bony plates termed osteoderms, and these give them a (partly flexible) carapace. Scutes can also occur in places where there are no underlying osteoderms. It is crucial for the remainder of this article that you remember and pay attention to the difference between osteoderms (bone) and scutes (keratin). Living armadillos are variable in terms of how much naked skin they possess, how hairy they are, and in how much of their skin away from the osteoderm-covered sections is decorated with scutes.

Caption: there aren’t that many glyptodont toys and figures, here are most (not all) of them. From left to right, these are variously by Mothman Republic, CollectA, Safari, Invicta (the three same-shaped ones), Colorata (the small one at the front), Schleich and K&M. A video about these figures is online here. Image: Darren Naish.
Given this diversity, what did glyptodonts look like when alive? Before we start thinking about the implications that extant armadillo appearance has for glyptodonts, we have to go to the fossils first. What direct evidence do we have to go on? As is so frequently the case when it comes to extinct animals, the answer is… not much! There is (to my knowledge) no reported glyptodont skin, no preserved keratinous covering of the osteoderms, no trackways or body impressions preserved in sediment that might be informative on this front, nor are there informative pieces of prehistoric rock art. Yes, there are glyptodont footprints, but they don’t reveal skin detail or anything like that (Aramayo et al. 2015). In addition, scarcely any of the scientific work we might be able to perform on glyptodont fossils has been done. Working out glyptodont life appearance just hasn’t been a topic that palaeontologists have invested much time in.

Caption: glyptodont tracks from the Late Pleistocene site of Pehuen Co paleoichnological site in Argentina. Dubbed Glyptodontichnus pehuencoensis by Aramayo et al. (2015), they show that digit II-IV made impressions with the substrate in the hand, and that all five toes fully contacted the ground. Glyptodonts have remarkably square, hoof-like unguals on most digits. Image: Aramayo et al. (2015).
The super armadillo model. So, what can we do? I put it that we can make some number of good or sensible guesses, and propose various arguments, extrapolations and speculations on glyptodont life appearance. My core assumption – and it’s hardly novel, since specialists on the group have already said much the same – is that glyptodonts are ‘super armadillos’: members of the group that took widespread armadillo traits to an extreme as they evolved a size and elaboration of armour that exceeded that of other members of the group. Connected to this assumption is the notion that glyptodonts hypertrophied their keratinization, which is a technical way of saying that they evolved a more extensive, more elaborate, more complete quantity of scute coverage than other armadillo lineages.

Caption: this ‘cross-sectional’ view of a Glyptodon (representing a view quite often shown in the literature) emphasizes the size of the shape of the cephalic osteoderm shield (the ‘skull cap’) and the dome-like form of the carapace. Osteoderms also covered much of the tail, there often being caudal rings and a more distal ‘tube’ of coossified osteoderms. This specimen is on show at the National Museum of Scotland, Edinburgh. Images: Darren Naish.
If glyptodonts were indeed hyper-keratinized, this has major impacts on life appearance. Let’s imagine those parts of the animal covered by osteoderms, namely the top of the skull, the greater part of the body (that part covered by the carapace, of course), and the tail. Glyptodonts would have been like living armadillos in that these osteoderms were covered in keratin scutes, but how thick these scutes were is – to my knowledge – unknown. But read on, we’ll get to this.
The data from osteoderms. The horny scutes of fossil armadillos in general might be unknown for the vast majority of taxa, but what do the osteoderms say on life appearance? The short answer is that they do provide at least some information.
Armadillo osteoderms are complex structures. For starters, the external or integumentary surface of the osteoderm is divided into a series of discrete ‘islands’, bizarrely termed ‘figures’ in the literature (I still have no idea why) separated by shallow grooves, sulci or channels (Klimeck et al. 2024). Foramina (pores or openings) of varying size occur across the surface, both over the ‘figures’ and within the sulci. A low number of relatively large, deep, vertically oriented foramina are located near the centre of the osteoderm. These are the glandular foramina and they connect to spherical or saccular cavities that house sweat and sebaceous glands (Krmpotic et al. 2009, 2015, Ciancio et al. 2019). A hair follicle is associated with the sebaceous gland in some groups, but not all.

Caption: a generic armadillo osteoderm, used to show key anatomical structures. Glyptodont osteoderms are highly modified relative to this basic plan. Image: Darren Naish.
More elongate, obliquely oriented, often (but not always) smaller foramina are distributed around the osteoderm margins. These are the marginal or piliferous foramina. They contain a hair follicle and either a small sebaceous or sweat gland (Krmpotic et al. 2015, Ciancio et al. 2019). They differ from glandular foramina in being connected internally by conduits, and in often being wider at their apertures than at their bases (Klimeck et al. 2024). Finally, osteoderms – being bones – have internal marrow cavities that are connected to various of the foramina mentioned here. How large and thick the marrow cavities are varies a lot within the group and the suggestion has been made that especially thick marrow cavities might correlate with adaptation to climate (Ciancio et al. 2019).
Because piliferous foramina support hairs, their number and distribution can give a rough guide to how hairy the carapace is or was. Extant armadillos with an especially hairy carapace have osteoderms peppered with abundant foramina of this sort. Among the best example of this is the Woolly long-nosed armadillo Dasypus pilosus where piliferous foramina are abundant and support numerous carapacial hairs (Castro et al. 2015).
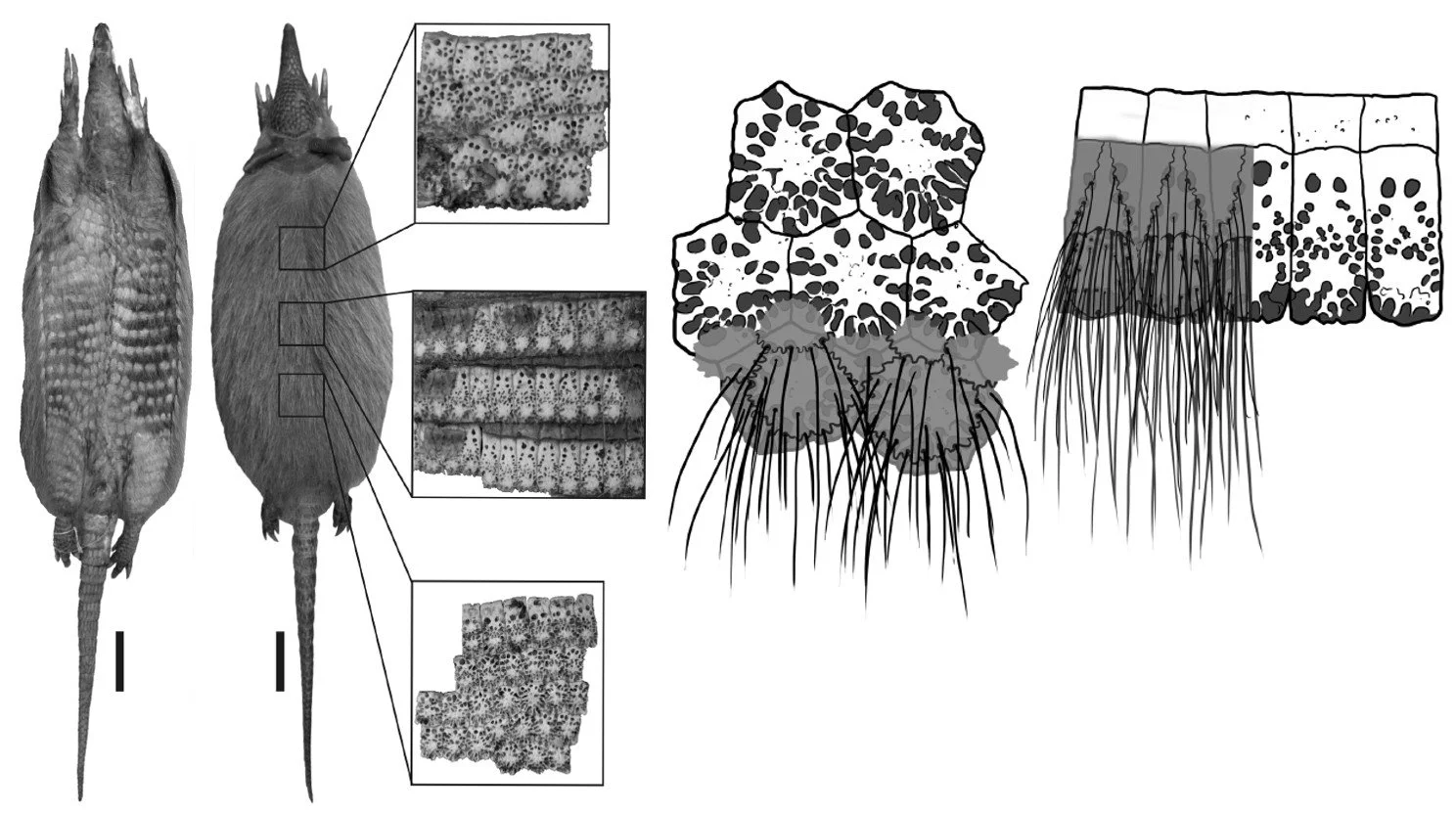
Caption: the number of piliferous foramina on the osteoderms do correlate with the number of hairs on the carapace, as illustrated here in these images of Woolly long-nosed armadillo Dasypus pilosus published by Castro et al. (2019). As is obvious, this visibly hairy species has a large number of these foramina on each osteoderm, typically between 15 and 30. I wrote about this species here. Images: Castro et al. (2019).
Some excellent studies of osteoderm architecture and histology and which include a good sampling of extinct armadilllos have been published, and the good news is that they provide great data on the external carapacial anatomy of these animals. Various euphractines and eutatines (see this article for those groups) were relatively hairy across parts of the carapace, at least, and so were stegotheriins (Ciancio et al. 2019), an unusual armadillo group I haven’t yet written about. A set of Eocene armadillos grouped together as the astegotheriins were not hairy on the carapace, and the suggestion has been made that an increased hairiness in geologically younger armadillos could be linked to cooling global temperatures (Ciancio et al. 2019).

Caption: at left, a 3D reconstruction of the osteoderm interior of Parutaetus oliveirai, an Eocene euphractine armadillo from Argentina, from Klimeck et al. (2024). The yellow blobs show the bone marrow cavities (bmc), the green ones correspond to the glandular cavitis (gc) and the red ones to the piliferous follicle cavites. Scale bar = 5 mm. At right, osteoderms of the fossil eutatines Utaetus and Parutaetus from Ciancio et al. (2019) in external view (at upper left), cross-sectional view (upper right), and as 3D interior reconstructions (the coloured images). I don’t include these images to show you the salient details, but to depict instead the high-quality work that now allows us to visualize osteoderm anatomy inside and out. Scale bars = 5 mm.
Scutes: thin or thick? A slightly surprising fact about the anatomy of living armadillos is that their scutes are sometimes extremely thin relative to the osteoderms, basically forming a thin ‘skin’ over the osteoderm surface. The osteoderms themselves are sometimes not thick, those of Nine-banded armadillo Dasypus novemcinctus being approximately 2 mm thick, and in such a case the surface keratin is about 200 μm (200 micrometres) thick (Chen et al. 2011). In that case, the scute is – according to the measurement data from Chen et al. (2011) – approximately one-tenth as thick as the osteoderm. That doesn’t seem consistent with published figures, however: in the one I show below, the scute is evidently very thin, but I can’t believe it’s one-tenth the thickness of the osteoderm.
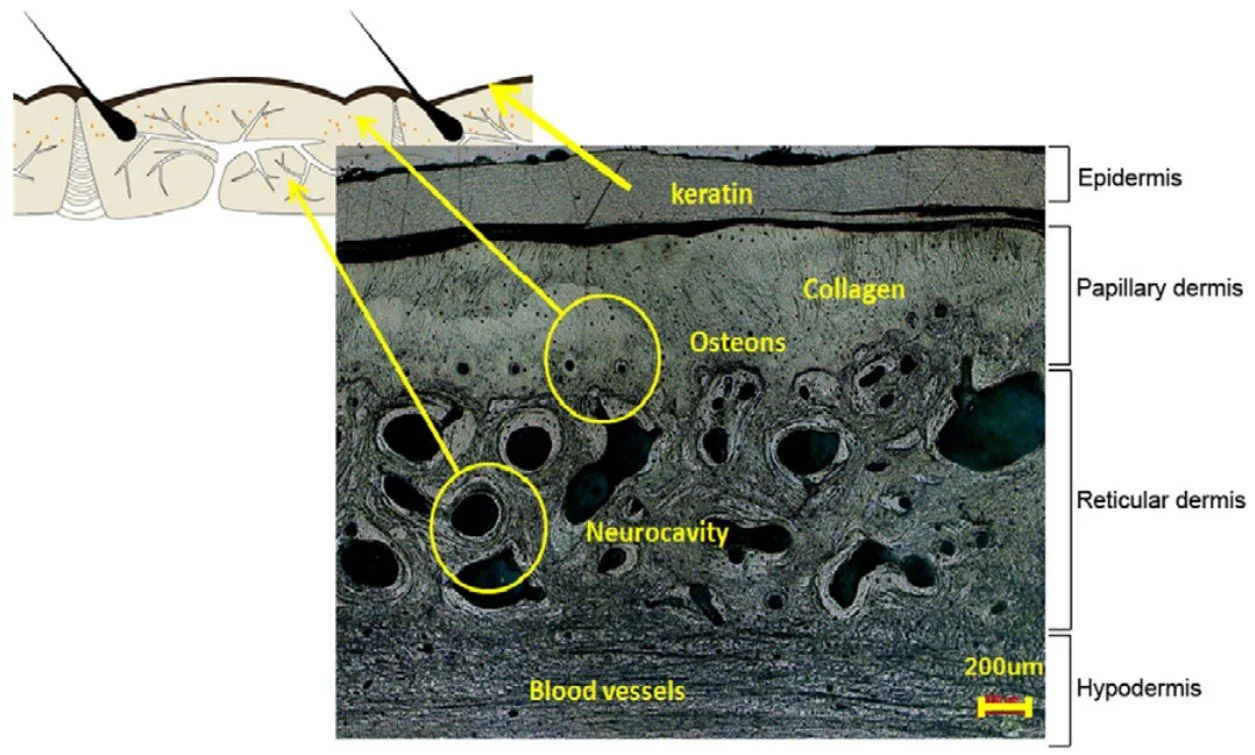
Caption: diagram and photo showing cross-section of Dasypus carapace from Chen et al. (2011), showing how thin the scutes are relative to the osteoderms. The photo, showing the internal microstructure of the osteoderm, doesn’t show the whole thickness of the structure but only its outermost layers. Image: Chen et al. (2011).
In any case, that data is from a dasypodid, and not from a sort of armadillo that might be a more applicable comparison to a glyptodont. That’s annoying, because things are evidently different among the armadillo groups. I don’t have measurement data, but photos of three-banded armadillo skeletons (see below) show that the scutes are (in places) approximately half as thick as the osteoderms. My hypothesis is that this is more applicable to the glyptodont condition. If you have ready access to armadillo remains, especially of euphractines or tolypeutines, please do test this and report the results. Fairy armadillos are probably not good as analogues since their burrowing lifestyle has clearly resulted in highly reduced osteoderms and scutes.

Caption: part of the lateral section of a Nine-banded armadillo carapace, the flexible, ‘banded’ section being at left. The scutes here (we’re seeing dried scutes, not osteoderms) possess between three and ten foramina, depending on position. This is not an obviously hairy species. Image: Darren Naish.
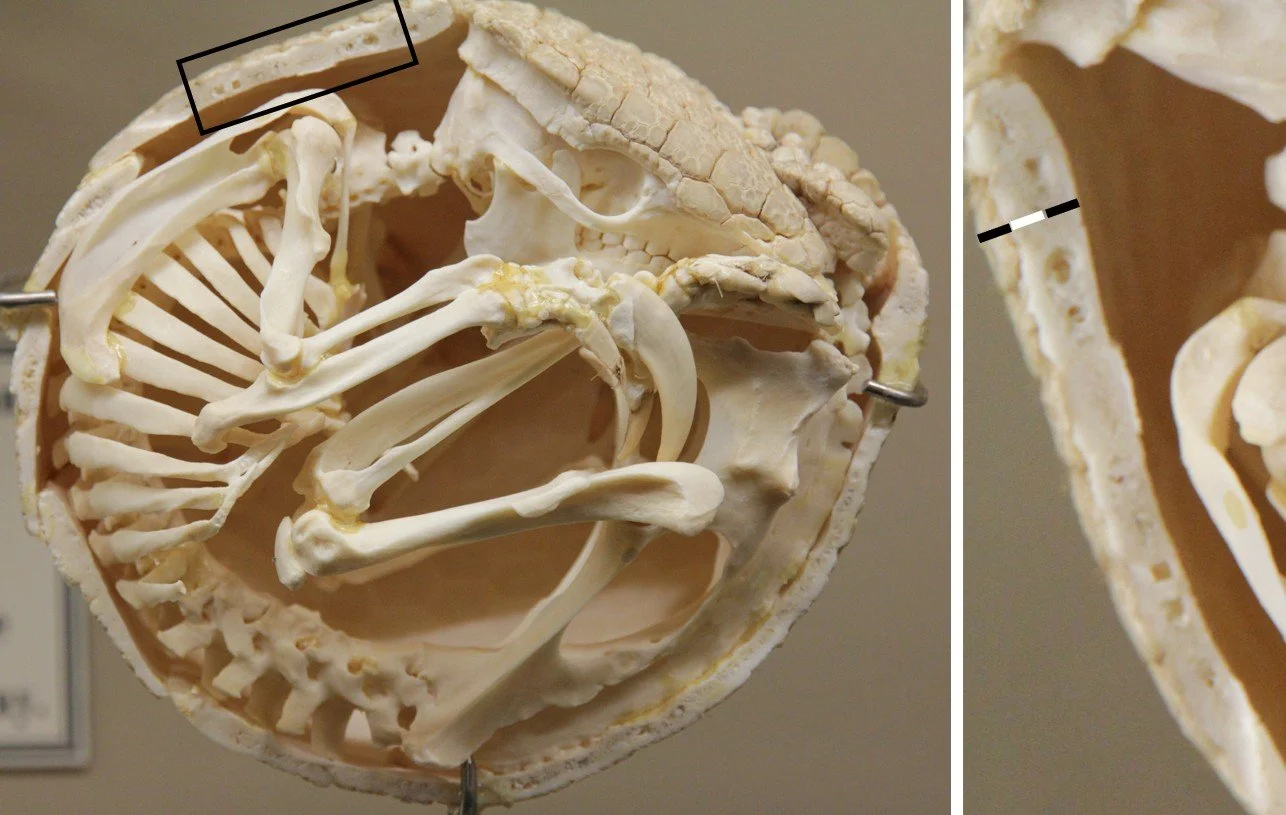
Caption: enrolled three-banded armadillo (Tolypeutes) skeleton, used here because it shows scute thickness relative to osteoderm thickness. I took the area shown in the dark rectangular and enlarged it; that’s the image at right. As you can see from the black-white-back bar I’ve added, the scutes in at least some places appear to be approximately 50% the thickness of the osteoderms. Image: Polyoutis, CC BY-SA 4.0 (original here).
Sooo… what about glyptodonts? Glyptodonts are mostly very weird compared to other armadillos. The osteoderms are unusual internally, containing large amounts of porous or spongy bone that make them thick and ‘inflated’ with substantial evidence for very active bone growth. Their external surfaces also look heavily remodelled (Hill 2006). This is all consistent with an active ‘function’ for the bones: it seems reasonable to speculate that they were able to repair themselves rapidly following combat-induced injury.
The osteoderms of big, adult glyptodonts are thick, as much as 3 cm (Luna et al. 2018). If what I said above about the scutes being one-third as thick as the osteoderms is right, we’re talking about a keratin thickness that would definitely affect the look of the animal in life. For one thing, the ornament present on the surfaces of the osteoderms would almost certainly be hidden from view: the scutes would have their own ornament, one you wouldn’t be able to predict from osteoderm anatomy (something that’s partly true of many living armadillos: the scute arrangement is often not a tight match for osteoderm placement). This means that at least some artistic reconstructions of glyptodonts are wrong, since they make it look as if the osteoderms were exposed in life and that the scute layer (over the skull cap and tail armour as well as the carapace) was as thin as it is in dasypodids. Keratin of reasonable thickness – arranged as hundreds of separate but close-fitting, polygonal mounds – would also have interesting and striking light-scattering qualities if backlit. In addition, those parts of the keratinous surface worn by abrasion against the ground, or by combat or some other repetitive behaviour, could have had a dulled, eroded or even notched or jagged appearance.
What else might the osteoderms say about life appearance? As a generalisation, glyptodont osteoderms have a rugose, irregular, even ‘pustulate’ or tuberculated external surface where ‘figures’ and sulci are indistinct and where small foramina are abundant (Hill 2006, Asakura et al. 2017, Perea et al. 2019). The giant, club-tailed form Doedicurus and its close relatives are unusual in having mostly smooth surfaces to their osteoderms (Hill 2006, Zurita et al. 2014).

Caption: three different carapacial osteoderms from Panochthus, all shown in external or integumentary view. Figures and sulci are more clearly visible in some (a) than others (c), and foramina large and small are visible in some (c) and might be absent in others (a). This sort of variation is present across the carapaces of most or maybe all glyptodonts. Scale bar = 20 mm. Image: Asakura et al. (2017).

Caption: carapacial osteoderms from Neuryurus, published by Perea et al. (2019). These images show (A) a large section from the front half of the carapace, specifically from a section on the upper part of the side; (B) close-up of osteoderms from the same section; (C) individual osteoderm (note the abundant foramina); (D) section from the edge of the carapace, the more rounded or conical osteoderms being from the carapacial edges. Images: Perea et al. (2019).
In contrast to other armadillos, the surface foramina of glyptodont osteoderms don’t group neatly into those of glandular and piliferous sort. Instead, you have a scattering of numerous small ones. In some glyptodonts, only some osteoderms – especially those at the edges of the carapace, but sometimes its dorsal midline too – have the occasional larger, more obliquely oriented foramen that should probably be identified as a piliferous one. Larger foramina of this sort are absent on cephalic and caudal osteoderms (Luna et al. 2018, Cuadrelli et al. 2019). Juvenile Glyptodon individuals have between three and seven larger (presumably piliferous) foramina on osteoderms that come from select portions of the carapace (Luna et al. 2018), but these are tiny, with diameters of 1 mm or less (Luna et al. 2018). Glyptotherium has piliferous foramina – as many as nine of them – on certain of its osteoderms but one or none on osteoderms from the side and rear portions of the carapace (Gillette et al. 2016). Doedicurus has large foramina – dubbed trans-osteodermal canals – on at least some of its osteoderms, those that have them possessing just one or two (Hill 2006).

Caption: these carapaces and (at left) select osteoderm segments reveal variation present across specimens of Glyptodon reticulatus from the Late Pleistocene and Holocene of southern South America, the A and B animals being adults, C being a juvenile. All the inset osteoderm segments are from the upper central portion of the carapace. Note the more prominent sculpting and higher number of foramina in C. In A and B, there are a low number of large, scattered foramina which may or may not have been hair follicles. Image: Cuadrelli et al. (2019).
My takehome from this anatomy is that the carapacial and caudal osteoderms of classic, giant glyptodonts – I’m talking here about Pliocene-Pleistocene-Holocene animals like Neuryurus, Glyptodon, Panochthus and Doedicurus – were largely hairless and covered by thick keratinous scutes. When hairs were present, they were small, sparsely distributed and mostly limited to the carapacial margins. This means that reconstructions where glyptodonts are given an especially hairy carapace – and they do exist – are not likely to be accurate.

Caption: a technically very good, interesting life reconstruction of the doedicurine glyptodont Eleutherocercus solidus, and showing it as a very hairy animal, notably across the carapace as well as across much of the rest of the body. Note also the hump above the pelvis with the distinct hair tuft. Image: Pedro Cuaranta, from Núñez-Blasco et al. (2021).

Caption: a low quality, crudely rendered image by myself, depicting various of the details up for discussion here. My Doedicurus here is hairy away from its osteoderms, but not that hairy, but lacks sufficient scutage (if you will) in the same areas. Its scute covering on the carapace does not correspond to the underlying osteoderm arrangement. When I first did the illustration (2015), I favoured the idea that the club might have short keratinous lumps at its terminus. I currently favour the idea of more formidable spines, hence the sketchy additions.
Incidentally, a few glyptodonts have a raised ‘dome’ on the carapace over the hips where the osteoderms are fused and especially big foramina are present. It’s been suggested that a gland of some sort was present here, in which case there might have been a tuft of long hairs or something of this sort (Núñez-Blasco et al. 2021).
Non-osteoderm-supported scutes. What about those parts of the body not covered by osteoderms? Living armadillos of several sorts possess scutes on parts of the skin where osteoderms are lacking, this being developed most prominently in three-banded armadillos. These animals have scutes across the cheek, around the eyes and sides of the snout, across the ears (both the inner and outer surfaces) and covering much of the limbs.
Caption: even certain small armadillos, like the three-banded armadillos, are ‘hyper-keratinized’, with non-osteoderm-supported scutes across the face, on both surfaces of the ears, and on the limbs all the way down to the claws. It is possible that glyptodonts were similarly extensively scute-covered. Image: Maurilbert, CC BY-SA 3.0 (original here).
The ‘super armadillo’ model requires that glyptodonts were like this, only more so. Scutes, according to this view, covered much of the side of the face, were confluent with the edges of the skull cap, covered the ears, and formed chainmail-like arrangements on the limbs all the way to the ends of the digits. Those scutes on the side of the face were presumably arranged in keeping with the topography of the underlying jugal flanges and other structures. If scutes possessed crests, keels, midline ridges or other features, these presumably exhibited an anatomically logical arrangement or flow, since that’s what they do in living armadillos, crocodylians, lizards and other animals with ornamented scales or scutes.
Hairiness or not. Armadillos of various sort are hairy, sometimes very so. The hairy armadillos (Chaetophractus) have an extensive amount of wiry hair growing across the underside, meaning that they have a proper pelt in this part of the body, and there are also hairs projecting from those scutes that cover the lower half of the carapace. A similar amount of hairiness is present in fairy armadillos, albeit not on the carapace. At the other extreme, the Giant armadillo Priodontes maximus is naked except for short carapacial hairs.

What does this mean for glyptodonts? It means that the floor is wide open in terms of how hairy we imagine them once we consider those areas away from the osteoderms. They could have been anywhere from wholly naked and lacking any sort of pelt all the way to fully furry. Even if largely naked, they may still have had vibrissae – whiskers – about the nose and mouth, and perhaps inside the ears. Maybe members of the group varied in hairiness according to the climates they were adapted to, since this was a big and diverse group that lived in numerous different habitats and climatic regimes.
The mace of Doedicurus. As ever, there’s still a lot to talk about. But one detail in particular has attracted special interest, and this is the remarkable tail club of Doedicurus. A striking arrangement of sockets small and large are arranged across the distal part of this structure, and the sad fact is that we don’t quite know what was going on here. Keratinous extensions of some sort were surely rooted within these sockets, but were they rounded lumps, hooked or straight spines, some combination of all of these things, or something else entirely, perhaps a semi-conjoined, macuahuitl-like structure?

Caption: a partial tail club of Doedicurus in a private collection, the human hands at far right giving you some idea of how massive and awesome this structure is. Large, mostly symmetrically arranged sockets are present on the sides, the dorsal and ventral surfaces, and on the posteriormost face. Images: Darren Naish.
All of these options are up for grabs and artists have explored most, but not all, of them. My own preference is for spines, partly because we know that glyptodonts with weaponized tails elsewhere in the family tree evolved tail spines of a different sort, and partly because I think they make most sense in terms of function. But this is speculative and I don’t think my feelings will be hurt if evidence demonstrates otherwise.

Caption: there have been several efforts to reconstruct the tail club of Doedicurus with massive spines, and two such reconstructions are shown here. The sockets show that there were a variable number of small structures on both the flattened dorsal and ventral surface, and much bigger ones around the edges. Both images are associated with museum displays but I haven’t been able to find their original sources; I’ll add source info when I find it.

Caption: we know that glyptodonts of several lineages evolved weaponized tails (there’s a study specifically on this by Victoria Arbor and Lindsay Zanno), and spiky osteoderms and scutes were present in at least some of these taxa. Caudal osteoderm shape in Glyptodon, shown here, indicates the presence of markedly pointed scutes (hence extrapolated black outline I’ve added). Image: John Hutchinson, from here.
A task unfinished. Here's where I have to end, with so many other things still wanting. Facial anatomy, what glyptodont body shape mean for mating behaviour and the anatomy of their genitals, their highly unusual hands and feet, and what we know (or think we know) of their sensory abilities. At this point, Tet Zoo ver 4 now hosts three or four articles on this incredible and exciting, geologically long-lived armadillo group, meaning that they’re reasonably well-served, more so than many other groups I’m interested in. I’d still like, at some point, to provide an overview of glyptodont diversity, since articles like the one you’ve just read don’t properly cover glyptodont phylogeny or taxonomy. Sigh, one day.
For previous Tet Zoo articles on armadillos and other xenarthrans, see…
Five things you didn’t know about armadillos, June 2007
What was that skull? (on glyptodonts), May 2008
I, Priodontes, the tatuasu, September 2008
Predation and Corpse-Eating in Armadillos, September 2010
The Fate of the Woolly Long-Nosed Armadillo of Peru, April 2024
Of Zaedyus, the Pichi, May 2025
Armadillo Empire, Part 1: of Euphractines and Eutatines, June 2025
Armadillo Empire, Part 2: Fairies, Tolypeutines, and Where Glyptodonts Go (July 2025)
Sloth World, 2025 (Part 1), October 2025
Caption: did I mention that I own a Doedicurus t-shirt?
Refs - -
Aramayo, S. A., Manera de Bianco, T., Bastianelli, N. V. & Melchor, R. N. 2015. Pehuen Co: updated taxonomic review of a late Pleistocene ichnological site in Argentina. Palaeogeography, Palaeoclimatology, Palaeoecology 439, 144-165.
Asakura, Y., Pareira, P. V. L. G. D. C., Oliveira, E. V. & Silva, J. L. L. D. 2017. Comparative paleohistology in osteoderms of Pleistocene Panochthus sp. Burmeister, 1886 and Neuryurus sp. Ameghino, 1889 (Xenarthra, Glyptodontidae). Comptes Rendus Palevol 16, 795-803.
Billet, G., Hautier, L., de Muizon, C. & Valentin, X. 2011. Oldest cingulate skulls provide congruence between morphological and molecular scenarios of armadillo evolution. Proceedings of the Royal Society B 278, 2791-2797.
Castro, M. C., Ciancio, M. R., Pacheco, V., Salas-Gismondi, R. M., Bostelmann, J. E. & Carlini, A. A. 2015. Reassessment of the hairy long-nosed armadillo “Dasypus” pilosus (Xenarthra, Dasypodidae) and revalidation of the genus Cryptophractus Fitzinger, 1856. Zootaxa 3947, 30-48.
Chen, I. H., Kiang, J. H., Correa, V., Lopez, M. I., Chen, P.-Y., McKittrick, J. & Meyers, M. A. 2011. Armadillo armor: mechanical testing and micro-structural evaluation. Journal of the Mechanical Behavior of Biomedical Materials 4, 713-722.
Ciancio, M. R., Krmpotic, C. M., Scarano, A. C. & Epele, M. B. 2019. Internal morphology of osteoderms of extinct armadillos and its relationship with environmental conditions. Journal of Mammalian Evolution 26, 71-83.
Cuadrelli, F., Zurita, A. E., Toriño, P., Miño-Boilini, Á. R., Rodríguez-Bualó, S., Perea, D. & Acuña Suárez, G. E. 2019. Late Pleistocene Glyptodontinae (Mammalia, Xenarthra, Glyptodontidae) from southern South America: a comprehensive review. Journal of Vertebrate Paleontology 38, e1525390.
Gilette, D. D., Carranza-Castañeda, Ó., White, R. S., Morgan, G. S., Thrasher, L. C., McCord, R. & McCullough, G. 2016. Ontogeny and sexual dimorphism of Glyptotherium texanum (Xenarthra, Cingulata) from the Pliocene and Pleistocene (Blancan and Irvingtonian NALMA) of Arizona, New Mexico, and Mexico. Journal of Mammalian Evolution 23, 133-154.
Hill, R. V. 2006. Comparative anatomy and histology of xenarthran osteoderms. Journal of Morphology 267, 1441-1460.
Klimeck, T. D. F., Ciancio, M. R., Sedor, F. A. & Kerber, L. 2024. Revealing the diversity of Paleogene cingulates from Brazil: a new species of Parutaetus (Euphractinae) in the Guabirotuba Formation (middle-late Eocene). Journal of Vertebrate Paleontology 44, e2403581.
Krmpotic, C. M., Ciancio, M. R., Barbeito, C. G., Mario, R. C. & Carlini, A. A. 2009. Osteoderm morphology in recent and fossil Euphractinae xenarthrans. Acta Zoologica 90, 339-351.
Krmpotic, C. M., Ciancio, M. R., Carlini, A. A, Castro, M. C., Scarano, A. C. & Barbeito, C. G. 2015. Comparative histology and ontogenetic change in the carapace of armadillos (Mammalia: Dasypodidae). Zoomorphology 134, 601-616.
Luna, C. A., Cerda, I. A., Zurita, A. E., Gonzalez, R., Prieto, M. C., Mothé, D. & Avilla, L. S. 2018. Distinguishing Quaternary glyptodontine cingulates in South America: how informative are juvenile specimens? Acta Palaeontologica Polonica 63, 159-170.
Mitchell, K. J., Scanferla, A., Soibelzon, E., Bonini, R., Ochoa, J. & Cooper, A. 2016. Ancient DNA from the extinct South American giant glyptodont Doedicurus sp. (Xenarthra: Glyptodontidae) reveals that glyptodonts evolved from Eocene armadillos. Molecular Ecology 25, 3499-3508.
Núñez-Blasco, A., Zurita, A. E., Miño-Boilini, A. R., Bonini,R. A. & Cuadrelli, F. 2021. The glyptodont Eleutherocercus solidus from the late Neogene of North-Western Argentina: morphology, chronology, and phylogeny. Acta Palaeontologica Polonica 66, 79-99.
Perea, D., Toriño, P. & Ghizzoni, M. 2019. First endoskeletal remains of Neuryurus (Xenarthra, Glyptodontidae), an emended diagnosis of the genus, and body mass estimations. Journal of Vertebrate Paleontology 39, e1668400.
Zurita, A. E., Taglioretti, M., de los Reyes, M., Oliva, C. & Scaglia, F. 2014. First Neogene skulls of Doedicurinae (Xenarthra, Glyptodontidae): morphology and phylogenetic implications, Historical Biology 28, 423-432.



