If you know anything about animals, you’ll know that hybridization – here meaning the crossing of species – is commonly reported among wild, free-living animals, and appears ubiquitous in some species pairs.

Caption: the Jurassic World universe has, over time, included more and and more hybrid dinosaurs, some very silly. 'Stegoceratops' – a genetic hybrid between Stegosaurus and some sort of ceratopsian (not necessarily Triceratops, maybe Nasutoceratops) – has been a mainstay of the games and expanded universe for a while. Image: Jurassic Park wiki (here).
We’ve known this for centuries, mostly because it’s obvious from the anatomy of the hybrid offspring but also because we’ve observed interspecies pairing – and its results – in action. Late 20th century advances in genetics have shown that hybridization is even more common than previously thought, and that whole populations, or sections of populations, have a hybrid ancestry.
We also know, thanks to centuries of keeping animals in captivity, that hybrids of most remarkable sort can occur when conditions allow, certain animal groups possessing behavioural, anatomical and genetic traits that permit an ‘anything goes’ approach to crossing (e.g., Allen & Short (1997) on equids, Arantes et al. (2020), Vilaça et al. (2021) on sea turtles, Káldy et al. (2020) on sturgeon x paddlefish hybrids). In cases, the species concerned are not especially closely related and have been going their separate evolutionary ways for 10, 20, or 30 or more million years.

Caption: hybrids that, by now, we all know and love. At left, Russian sturgeon Acipenser gueldenstaedtii (a) and American paddlefish Polyodon spathula (d), and hybrids (sturddlefish) between the two. These are captive hybrids and couldn’t have come about in the wild. At right, juvenile hybrid sea turtle captured off Florida in 2016. It looks like a Green turtle Chelonia mydas but has a few traits characteristic of Loggerhead Caretta caretta. These species belong to lineages that separated between 35 and 46 million years ago [UPDATE: see comments. Those claims of very old divergences in extant sea turtles are probably very wrong]. Images: Káldy et al. (2020); Shamblin et al. (2018).
Because the hybrid animals that have received the greatest amount of discussion are big mammals – like canids, bears, zebras and cetaceans – you might have the impression that hybridization in the wild is a mostly mammals, perhaps even mammals-only, thing. It totally isn’t. We know of cases in which related bird species hybridize in the wild, examples including ducks, grouse, gulls, hummingbirds and corvids. Among non-bird reptiles, wild hybrids are known among sea turtles (Arantes et al. 2020, Vilaça et al. 2021), Galapagos tortoises (Miller et al. 2017), crocodiles (Milián-García et al. 2011, 2015) and numerous lizards (Rassman et al. 1997, Reeder et al. 2002, Jančúchová-Lásková et al. 2015). In amphibians, certain western Palaearctic water frogs (Spolsky & Uzzell 1986) and ambystomatid salamanders famously hybridize (Spolsky et al. 1992). Wild-living fish hybrids are well known among salmon and carp.

Captions: if you look at gulls a lot (the white-headed Larus species especially), you’ll know that hybrids are moderately common in some populations, and often difficult to identify precisely. At left, a gull photographed in Lisbon in 2014 and which seems to combine traits of Lesser black-backed L. fuscus and Herring gulls L. argentatus. At right, an Olympic or Puget Sound gull photographed in Oregon in 2022. This sort of gull appears to be a hybrid between Western L. occidentalis and Glaucous-winged gulls L. glaucescens. Images: Darren Naish.
Why animals hybridize. Why animals hybridize is a complex question with several answers. Certain animals recognize the vocal, acoustic and visual signals of close relatives as ‘similar enough’ to those of their own species that they function as appropriate stimuli when the time is right. In those cases, hybridization might be described as opportunistic.
Incidentally, an idea popular in the dinosaur literature – that the extravagant horns, frills, crests and so on of dinosaurs helped the species recognise their own and avoid mating with the members of other species – is surely mostly erroneous (Hone & Naish 2013). They don’t function that way in living species. It was more likely a combination of factors, involving pigmentation, behaviour and olfactory and vocal signals, that ‘controlled’ courtship and mating events.
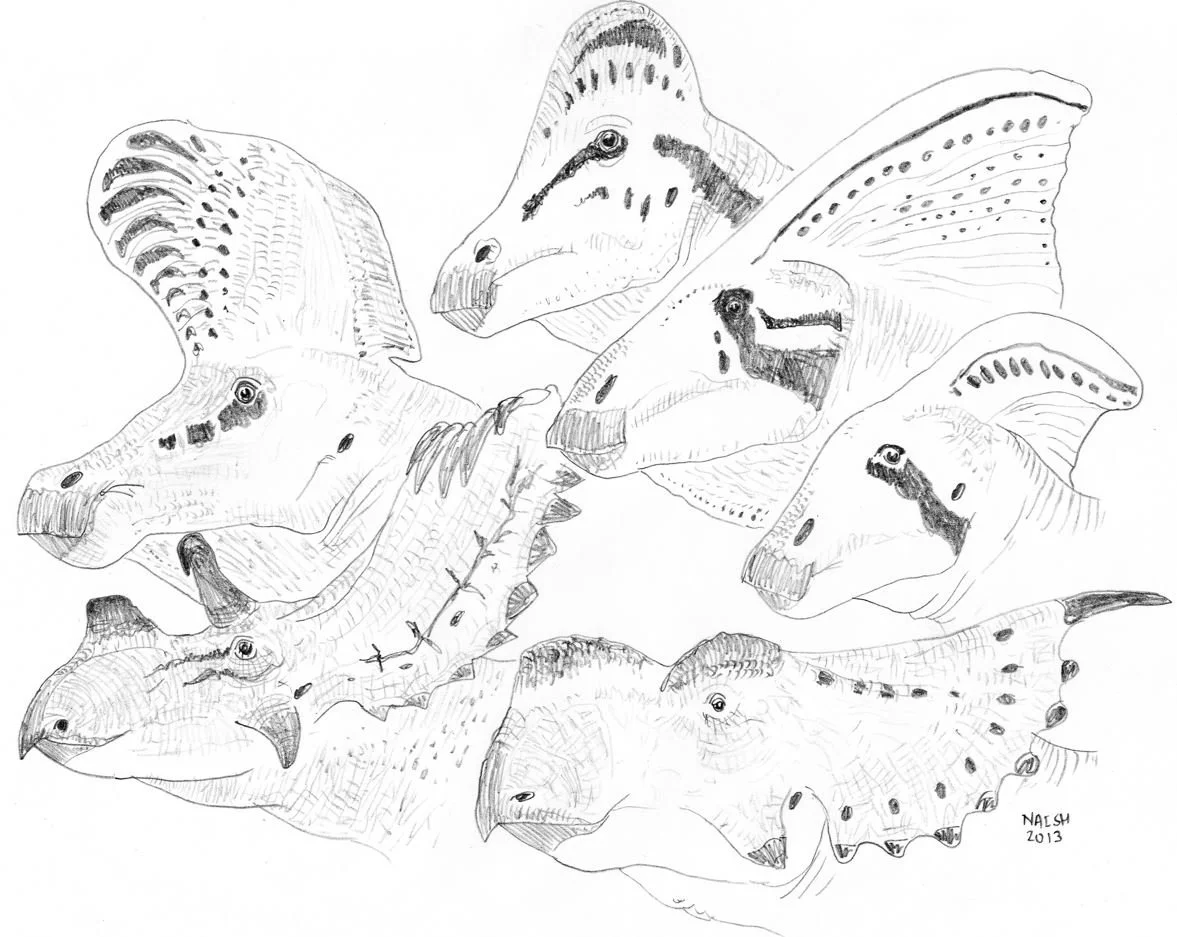
Caption: there are numerous dinosaur communities where some or several close relatives lived alongside one another; the animals shown here weren’t all sympatric but are representatives of certain of the relevant groups (lambeosaurine hadrosaurs and both chasmosaurine and centrosaurine ceratopsids). These animals often exhibit extravagant structures but – even so – we might speculate that hybridization occurred here and there. This illustration is looking very dated. Image: Darren Naish.
I digress. In some cases, it’s hypothesized that hybridization occurs because individuals fail to find mates of their own species. From an evolutionary perspective, a ‘bad mating’ is better than no mating at all: there’s an advantage in using opportunities to pass on genes, species boundaries be damned. Species that are rare (due to human hunting, habitat loss or deterioration and so on) may therefore have little option other than to hybridize. At least some of the hybridization we see in the modern world might, then, not have occurred – or, at least, not have occurred at regular frequency – in the geological past. Having said that, a world devoid of human pressures could still result in situations where species were forced to hybridize with related species, and we would do well to keep this in mind when looking at fossils from ecosystems thought to have been under environmental stress.
A second possible impact of humans on hybridization might run the other way: that is, that the declines we’ve caused to species and populations might reduce the opportunity for hybridization, since we’ve lowered the number of cases where species are sympatric and thereby lowered the opportunities for hybridization. The fossil record shows, in general, that sympatry between related species was more common in the past than it is today. Indeed, many ancient ecosystems look packed in terms of related species relative to modern ones. Don’t forget that the world today is depauperate when it comes to megafaunal diversity and distribution.

Caption: it’s well known that Panthera species can hybridize, with ligers (Panthera leo x P. tigris) being probably the most familiar. Ligers have been known to science since the late 1700s and can be (relatively) easily created in captivity. Notably, the two parent species here have, in the past, occurred sympatrically and there are unconfirmed accounts of their hybrids existing in the wild. Image: Camphora, public domain (original here).
Are hybrids really ‘common’? Not in birds. Hybridization in some animal groups is, statistically speaking, rare and statements implying that it’s common or ubiquitous need to be seen in context. Yes, hybrids between species x and species y might often get reported (indeed, hybrids are deemed so interesting that they’re almost certainly over-reported), but they’re rare in terms of frequency.
Ernst Mayr, one of the most influential and famous figures in our understanding of bird evolution, suggested on the basis of museum specimens that hybrids occurred on the order of 1 in 60,000, meaning that about 0.001% of individuals were hybrids (Mayr 1963, p. 114). More recently, Justyn et al. (2020) used eBird data for the United States collected between 2010 and 2018 and found that hybrids accounted for 0.064% of records. That’s a higher percentage than Mayr’s, this perhaps explained by differing views on species boundaries (the gulls and ducks that modern workers regard as distinct species were not treated as such by Mayr). When very ‘hybrid-heavy’ species were excluded (like Mallard Anas platyrhynchos and Mexican duck A. diazi), hybridization frequency was 0.009% (Justyn et al. 2020).

Caption: two ducks that engage in a lot of hybridizing. Mexican duck at left, Mallard at right. Images: ALAN SCHMIERER, CC0 (original here); Darren Naish.
In addition, nearly 86% of all bird hybrids in the dataset compiled by Justyn et al. (2020) involved just ten species pairs, all of which were ducks, geese, gulls and chickadees. In fact, nearly 83% of all US hybridization events involved ducks and gulls alone. Huge swathes of the avian tree of life (at least, based on US data) have no meaningful hybridization going on at all (Justyn et al. 2020).
Crocodylians, turtles and squamates. The take-home from this perspective is that hybridization is rare overall, is aberrant and remarkable across big swathes of the avian tree and is only ‘expected’ or ‘usual’ if you’re dealing with a very particular, very small set of unusual species. It’s not clear how applicable this observation is when it comes to other animal groups though, and it should be obvious from the title of this article that I’m especially interested here in possible hybridization among non-bird dinosaurs. In view of that, the animals most relevant after birds are crocodylians, turtles and squamates. How common, or rare, is hybridization in those animals?
For crocodylians, cases of hybridization among wild animals have been well recorded between certain species pairs, namely American crocodiles Crocodylus acutus and Morelet’s crocodile C. moreletii in the Mexican Caribbean (e.g., Machkour-M’Rabet et al. 2009), and Cuban crocodiles C. rhombifer and a population historically (but probably incorrectly) identified as American crocodiles in the Zapata Swamp of Cuba (e.g., Milián-García et al. 2011, 2015). In the Zapata Swamp case, as many as 49% of the wild-living individuals had a hybrid ancestry (Milián-García et al. 2015) and a repeated claim made in the relevant publications is that hybridization among crocodile species is common.

Caption: Cuban crocodile, a species that has hybridized in the wild with the related crocodile species. Whole populations in parts of Cuba appear to have a hybrid ancestry but human intervention may have a role in contributing to this. The Cuban crocodile is a fantastic animal, highly capable on land, good at grabbing animals from overhead branches (by leaping from water), and with great pigmentation. Image: Zanbog, CC BY-SA 2.0 (original here).
However… we’re referring here to declining, relictual populations of species whose ranges have been substantially reduced relative to the historical norm. Indeed, it’s been suggested that this hybridization might have been “enhanced by anthropogenic pressures”, to quote Cuban crocodile expert Yoamel Milián-García. Is hybridization in crocodiles really ‘common’? I would challenge that contention.
Among lizards, hybridization in the wild is known among around 70 species, including assorted iguanas (Rassman et al. 1997, Vuillaume et al. 2015, Moss et al. 2018), fence lizards, anoles and other iguanians (Jančúchová-Lásková et al. 2015), whiptails (Reeder et al. 2002), certain snakes, lacertids and several gecko groups (Jančúchová-Lásková et al. 2015). Some whiptail species appear to owe their origins to hybridization.
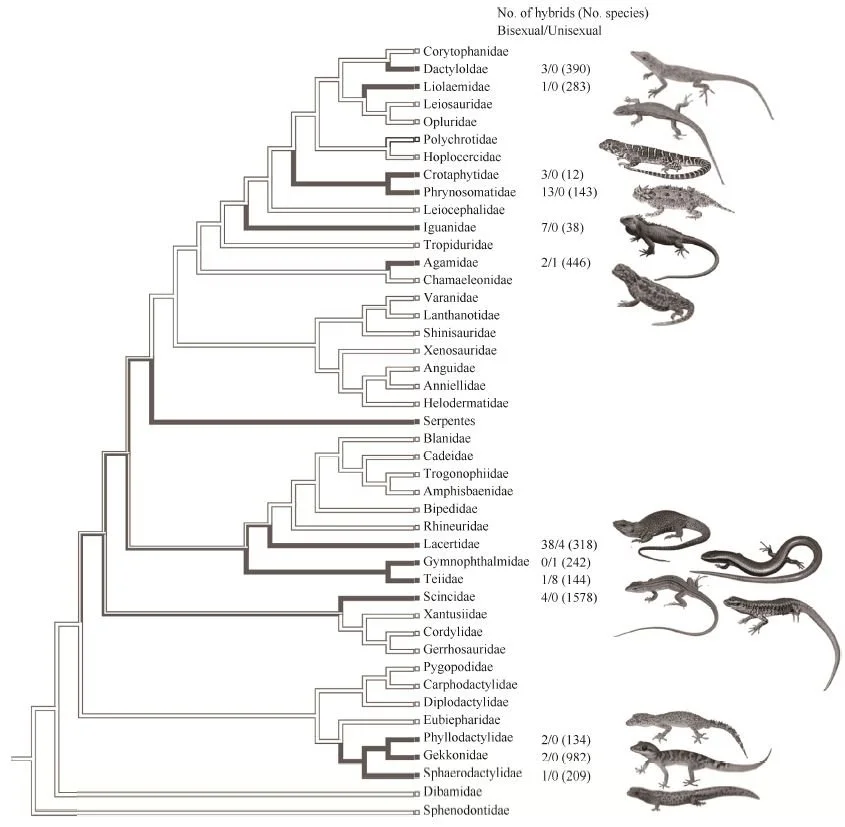
Caption: very useful phylogenetic tree from Jančúchová-Lásková et al. (2015) showing prevalence of hybridization within lizards. It's widespread enough that we should regard it as ubiquitous across the group, but note that there are numerous groups where it basically hasn't been reported.
This indicates that an ability to hybridize is widespread among lizards, but “[r]eliable records of hybridization are scarce” (Jančúchová-Lásková et al. 2015, p. 169) and, again, the numbers of species we’re dealing with are low relative to the number of lizards there are in total. In addition, at least some examples (those involving Iguana species) are due to human introduction. Yes, hybridization occurs between some species pairs, but it shouldn’t be regarded as widespread across the animals as a whole, nor ‘normal’ or ‘ubiquitous’.
Very similar things could be said about turtles (which now appear to be on the archosaur lineage and thus closer to dinosaurs than lizards). Assorted rare hybridizations have been reported from the wild (Miller et al. 2017, Arantes et al. 2020) and at least some of these, as well as some from captivity (Buskirk et al. 2005), are consistent with the ‘anything goes’ tendency noted earlier. Again, some and maybe even most or all of the recorded wild hybridizations are linked to human-caused factors.
In quest of hybrid (non-bird) dinosaurs. Here we come to the raison d’âtre of this article. If hybridization occurs among animals of so many sorts today, we absolutely can and should assume that it occurred among the animals of the past. We could discuss this issue as it pertains to fossil hominins, elephants, crocodylians or frogs, but we’re here because of the non-bird dinosaurs.

Caption: the very famous Triceratops specimen AMNH 5116, on show at the American Museum of Natural History in New York. It's a really famous Triceratops but also a deeply odd one. Image: Kabacchi, CC BY 2.0 (original here).
My primary contention (or assumption) is that hybrid non-bird dinosaurs did exist as live animals. Based, however, on the points we’ve already seen, we should be working on the assumption that hybridization was rare, if not extremely so. It remains possible that there were certain populations or species – like those duck and gull species noted above – where hybridization was frequent.
While we don’t have a firm idea of which species pairs might have been ‘gull- or duck-like’ in their propensity to hybridize, an argument can be made that we can predict which species pairs were most likely to do so. Obviously, we might start with closely related, anatomically similar, sympatric animals. Late Cretaceous lambeosaurines and chasmosaurines come to mind, as do Morrison Formation diplodocids, Wessex Formation iguanodontians, and the Late Cretaceous east Asian ceratopsians grouped together as protoceratopsids or bagaceratopids (yes, different group name endings there), for starters.
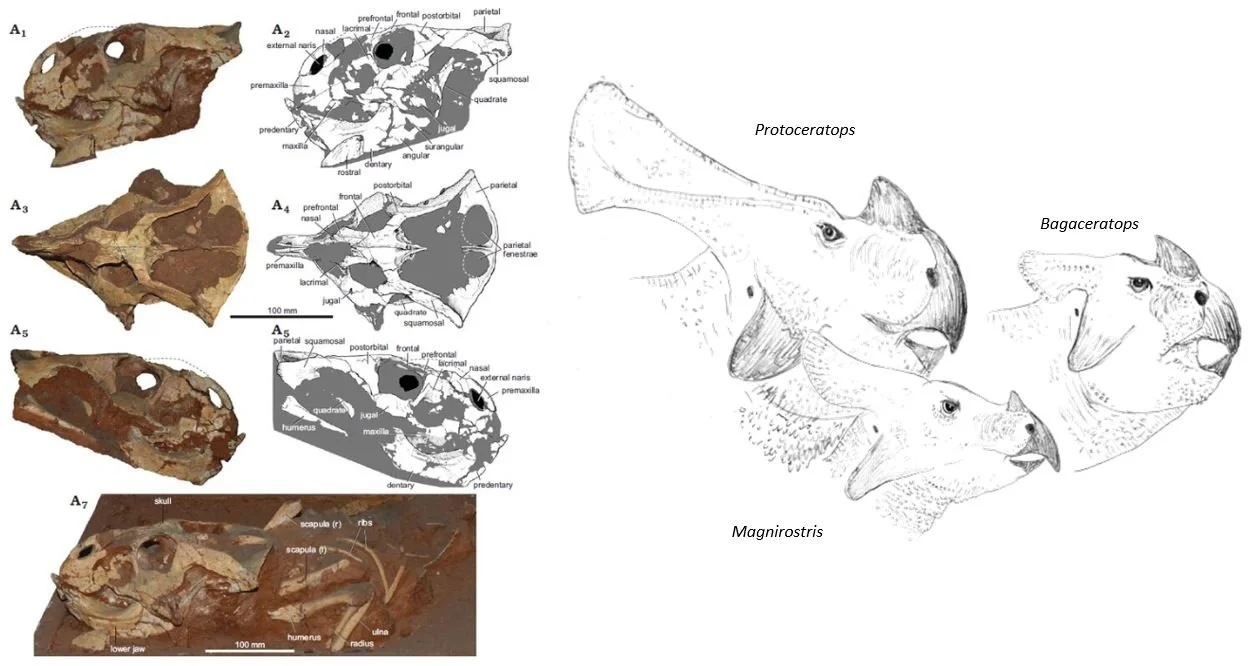
Caption: at left, the ceratopsian specimen MPC-D 100/551B from Ömnögovi, Mongolia, identified by Czepiński (2020) as ‘cf Bagaceratops sp’ and suggested to perhaps be a hybrid. At right: opinions differ among experts as to how many taxa should be recognized among Late Cretaceous east Asian ceratopsians (Magnirostris, shown here, is generally thought today to be synonymous with Bagaceratops). If any of these animals were sympatric, it is at least plausible that hybridization occurred. Images: Czepiński (2020); Darren Naish.
A suggestion already exists that a Bagaceratops-like ceratopsian from the Gobi Desert might be a hybrid between Protoceratops andrewsi and B. rozhdestvenskyi (Czepiński 2020) but alternative explanations for its anatomy – that it represents a transitional evolutionary form between the two or is simply a new species – is on the cards too. Various Triceratops specimens look anatomically ‘intermediate’ between the older T. horridus and the younger, shorter-snouted T. prorsus (Scannella et al. 2014). The favoured explanation is that these are, again, evolutionary intermediates, consistent with the view that the younger species evolved directly from the older one, and likewise for other ceratopsids that look like ‘intermediates’ (Fowler & Freedman Fowler 2020). Comments have occasionally been made about the Triceratops on show in New York (AMNH 5116), since (while heavily reconstructed in the frill especially) it looks something like an intermediate between Triceratops and Torosaurus, this at least raising the possibility that it might be a hybrid between the two.
Caption: several Triceratops specimens (like UCMP 113697 and MOR 3027, shown here in the middle part of the tree) were found by Scannella et al. (2014) to be 'intermediates' between the two recognized species. Within their hypothesis of anagenetic evolution, such animals must have existed. But the possibility that some of these specimens might be hybrids is on the cards. Image: Scannella et al. (2014).
Caption: the famous Yoshi’s Trike, MOR 3027, at left, on show at the Museum of the Rockies, Montana. This animal is anatomically intermediate between the Triceratops species T. prorsus and T. horridus... could it be a hybrid? That’s a fun idea, since its ridiculous supraorbital horns could then be imagined (I said imagined) as a consequence of hybrid vigour. Image: WernerG2011, CC BY-SA 2.0 (original here).
Another point: even if we do find two species that might have engaged in hybridization, we have no way of knowing how typical they were of their broader groups. Stated another way, just because two species in a group of 50 regularly hybridize, it doesn’t follow that regular hybridization should be assumed for the remaining 48 within the group.
Can we test for hybridization in fossil dinosaurs? Even if hybridization did occur, regularly or not, the big question is… will we ever be able to do more than speculate about it? Will we ever be able to demonstrate it? We don’t exactly have large datasets for the majority of fossil dinosaur species. Most are known from singletons.
In living animals, we can spot hybrids because they combine physical traits belonging to both parent species. And if we measure the physical traits of hybrid individuals, we often find that they’re anatomically intermediate between their parent species. Could we determine, via morphometrics, that a given fossil dinosaur specimen is a hybrid? A sample size of between 20 and 30 individuals is considered exceptionally good for fossil dinosaurs; we’re virtually always well short of the few hundred you need to be confident about the distribution of shape and size within a population. If we do find a specimen that’s unusual in morphometric terms (that is, it’s intermediate between individuals of species x and species y, or has proportions that make it partly overlap either or both x and y), our default and most conservative hypothesis is that we’ve found a new species.

Caption: Rackelhuhn, a naturally occurring and well known European hybrid between Western capercaillie and Black grouse. Image: F. C. Robiller, CC BY-SA 3.0 (original here).
One group of dinosaurs is represented by numerically sufficient samples to test for hybrids, and here I’m referring to Pleistocene and Holocene birds. Based on the prevalence of hybrids between three grouse species pairs – the Western capercaillie Tetrao urogallus and Black grouse T. tetrix, Western capercaillie and Willow ptarmigan Lagopus lagopus and Black grouse and Willow ptarmigan – Bocheński & Tomek (2000) looked at the bones of taxiderm hybrid specimens to see if their hybrid nature could be detected via osteology alone. They found that some animals could be identified as hybrids based on size and anatomical detail but that not all could, and they concluded that “it is probably only a matter of time to prove the presence of galliform hybrids in fossil and archaeological materials” (Bocheński & Tomek 2000, p. 698). This, and cases like it, are not exactly applicable to extinct species, however, since this is a case where we know that hybridization occurs, and have then gone looking for it in fossils. If we only had fossil grouse to go on, would we know that specimens with ‘intermediate’ measurements and details represent hybrids? That’s impossible to answer, but the answer is probably no.
Caption: Bocheński & Tomek (2000) showed that hybrids between extant galliform species could be identified osteologically, if you know exactly what to look for. As shown in these and other diagrams, known hybrids possess small osteological details in the sternum, coracoid, humerus and elsewhere in the skeleton that are intermediate between the conditions of the two parent species. Images: Bocheński & Tomek (2000).
Despite all of these caveats, excuses and handwringing: yes, there are specimens that look like they might be hybrids. The bad news is that we have no real way of testing, let alone confirming, a potential hybrid origin. In addition, the statistical rarity of hybridization relative to other potential explanations for an unusual anatomical configuration – in particular the possibility that the fossil concerned represents a new taxon of conventional, non-hybrid origin, or an evolutionary intermediate – renders any suggestions of a hybrid origin speculative in the extreme.

Caption: palaeoartists have indulged in a modicum of speculation on the sorts of hybrids that might have existed in some Mesozoic dinosaur communities. This hypothetical ‘Toroceratops’ represents a Triceratops x Torosaurus hybrid. Image: FALC Paleoart, used with permission.
‘Impossible’ hybrids that are not impossible. I want to end this article by playing with even more speculation, because where else can you do this if not on a blog. All the ideas about the possible presence of hybridization discussed above concern closely related taxa. But more exciting, and even more speculative, is the idea – again, inspired by hybrids in the modern world – that species only distantly related might hybridize too.
It’s highly unlikely that species from far-flung branches on the dinosaur family tree might have got it on with one another (no, for example, thyreophoran x ceratopsian crosses), but… based on hypothesized hybridization events that seem to exist across lineages within deer (Pitra et al. 2004), turtles (Buskirk et al. 2005, Arantes et al. 2020) and such birds as woodpeckers (Fuchs et al. 2013), it’s possible that extinct dinosaurs that were part of the same ‘family-level’ clade, but were not necessarily all that close, could still produce viable crosses. So, imagine a cross between a Parasaurolophus-like lambeosaurine and a Corythosaurus-like one, or between a Chasmosaurus-like chasmosaurine and something like Anchiceratops or Regaliceratops.
Caption: anatomy and genetics indicate that several extant animal species or groups probably do owe their origins to hybridization between species that are not especially closely related. As a consequence they do look weird and have often been hard to place in phylogenetic terms. Examples include the Milu or Père david’s deer Cervus davidianus (captive individual at left), and the campephiline woodpeckers (Crimson-crested woodpecker Campephilus melanoleucas shown here). I wrote about the ancestry of the Milu here. Images: Darren Naish; Bernard Dupont, CC BY-SA 2.0 (original here).
Even more extreme crosses – occurring between lineages distinct for over 20 or 30 million years – are also at least plausible based on what some living animals have done. Of course, we can’t say anything about the real probability of this given our lack of knowledge on their genetics. But it does mean that (arguably) ridiculous things are conceivable, like hybrids between centrosaurine and chasmosaurine horned dinosaurs, between apatosaurine and diplodocine sauropods, between microraptorines and velociraptorines within maniraptorans, or, shock horror, between baryonychines and spinosaurines within megalosauroids.

Caption: another hypothetical Late Cretaceous hybrid, this time one between members of the Parasaurolophus and Lambeosaurus lineages within lambeosaurines. Would such an animal succeed in winning mates itself? Would it be a loner, would it live with one or either parent species, and would its unique anatomy be reflected in unique acoustics? Questions we will likely never have answered… should such an animal have existed. Image: Hodari Nundu, used with permission.
To sum up this article in the most concise way possible… Could hybrid (non-bird) dinosaurs exists: yes, of course, they surely did, but they were likely vanishingly rare, so rare that they’re unlikely to be represented in the current palaeontological sample. If they are represented, it will be among species known from a good number of individuals. Can we ever identify hybrids, even among such species? Based on our current knowledge and analytical techniques, almost certainly not, but we should remain open-minded to the possibility that they might be both findable and identifiable.
For previous articles on Mesozoic dinosaur biology relevant to the topics covered here, see…
Could We Domesticate (Non-Bird) Dinosaurs?, August 2018
Postcranial Palaeoneurology and the Lifestyles of Pterosaurs, August 2018
Alternative Timeline Dinosaurs, the View From 2019 (Part 3): the Dinosauroid and its Chums, December 2019
Pete Dunne and Kevin T. Karlson’s Gulls Simplified: A Comparative Approach to Identification, October 2021
A brain for Baryonyx: using CT-scanning to examine British spinosaurid brains, February 2023
T. rex the Tool Maker -- Testing Controversial Claims About Dinosaur Smarts, April 2024
Refs - -
Allen, W. R. & Short, R. V. 1997. Interspecific and extraspecific pregnancies in equids: anything goes. The Journal of Heredity 88, 384-392.
Bocheński, Z. & Tomek, T. 2000. Identification of bones of galliform hybrids. Journal of Archaeological Science 27, 691-698.
Buskirk, J. R., Parham, J. F. & Feldman, C. R. 2005. On the hybridisation between two distantly related Asian turtles (Testudines: Sacalia × Mauremys). Salamandra 41, 21-26.
Fuchs, J., Pons, J.-M., Liu, L., Ericson, P. G. P., Couloux, A. & Pasquet, E. 2013. A multi-locus phylogeny suggests an ancient hybridization event between Campephilus and melanerpine woodpeckers (Aves: Picidae). Molecular Phylogenetics and Evolution 67, 578-588.
Hone, D. W. E. & Naish, D. 2013. The ‘species recognition hypothesis’ does not explain the presence and evolution of exaggerated structures in non-avialan dinosaurs. Journal of Zoology 290, 172-180.
Jančúchová-Lásková, J., Landova, E. & Frynta, D. 2015. Are genetically distinct lizard species able to hybridize? A review. Current Zoology 61, 155-180.
Justyn, N. M., Callaghan, C. T. & Hill, G. E. 2020. Birds rarely hybridize: a citizen science approach to estimating rates of hybridization in the wild. Evolution 74, 1216-1223.
Machkour-M’Rabet, S., Henaut, Y., Charruau, P., Gevrey, M., Winterton, P. & Legal, L. 2009. Between introgression events and fragmentation, islands are the last refuge for the American crocodile in Caribbean Mexico. Marine Biology156, 1321-1333.
Mayr, E. 1963. Animal Species and Evolution. Harvard University Press, Cambridge, MA.
Milián-García, Y., Venegas-Anaya, M., Frías-Soler, R., Crawford, A. J., Ramos-Targarona, R., Rodríguez-Soberón, R., Alonso-Tabet, T. J., Sanjur, O, Espinosa-López, G. & Beringham, E. 2011. Evolutionary history of Cuban Crocodiles Crocodylus rhombifer and Crocodylus acutus inferred from multilocus markers. Journal of Experimental Zoology 315, 358-375.
Moss, J. B., Welch, M. E., Burton, F. J., Vallee, M. V., Houlcroft, E. W., Laaser, T. & Gerber, G. P. 2018. First evidence for crossbreeding between invasive Iguana iguana and the native rock iguana (Genus Cyclura) on Little Cayman Island. Biological Invasions 20, 817-823.
Pitra, C., Fickel, J., Meijaard, E. & Groves, P. C. 2004. Evolution and phylogeny of old world deer. Molecular Phylogenetics and Evolution 33, 880-895.
Rassman, K., Trillmich, F. & Tautz, D. 1997. Hybridisation between the Galapagos land and marine iguana (Conolophus subcristatus and Amblyrhynchus cristatus) on Plaza Sur. Journal of Zoology 242, 729-739.
Shamblin, B. M., Mansfield, K. L., Seney, E. E., Long, C. A., Bagley, D. A. & Nairn, C. J. 2018. Brazilian origin of a neritic juvenile hybrid loggerhead x green turtle foraging in Florida. Marine Turtle Newsletter 155, 4-7.
Spolsky, C., Phillips, C. A. & Uzzell, T. 1992. Gynogenetic reproduction in hybrid mole salamanders (genus Ambystoma). Evolution 46, 1935-1944.
Spolsky, C. & Uzzell, T. 1986. Evolutionary history of the hybridogenetic hybrid frog Rana esculenta as deduced from mtNDA analyses. Molecular Biology and Evolution 3, 44-56.



