I’ve said before that proboscideans – the familiar group of placental mammals that includes living elephants and their many fossil relatives – have never been well served here at Tet Zoo…

Caption: most of us are fascinated by mammalian megafauna. That in mind, it’s difficult to express how incredible our planet was during the years of the Pleistocene. Here, a pair of the incredible scimitar-toothed cat Homotherium aim to isolate and kill a Woolly mammoth calf, a piece of behaviour attested by fossil evidence. Image: Hodari Nundu, used with permission.
For no specific reason at all, they’re just one of those groups of animals that I’ve never gotten round to writing all that much about. I aim to change this in future. Today, I want to share brief thought on one specific and very famous, and comparatively well understood, extinct proboscidean: the Woolly mammoth Mammuthus primigenius…
Mammoth basics. For starters, it’s always important to remind people that ‘the Woolly mammoth’ shouldn’t be referred to as ‘the mammoth’ given that mammoths include a cluster of species, varying in size, form, adaptation, habitat and geographical range. The Woolly mammoth – a cold-adapted, shaggy-coated species of northern Eurasia and North America – is among the most atypical and specialized of mammoths, and not an exemplar of what mammoths as a whole were like.

Caption: one of the UK’s best Woolly mammoth skulls, that of the Ilford (or Ilford Lane) mammoth, on show at the Natural History Museum in London. At left is how it looked in 2022; at right, its new location as of 2025. This provides a nice illustration of how the strong inward-and-upward curvature of the tusks is obvious in front view, and less clear from the side. The Ilford mammoth was collected in 1864; Ilford Lane is in Redbridge in London’s north-east. Images: Darren Naish.
It’s also important to remind people that mammoths are not ‘distant cousins’ or ‘relatives’ of elephants: they are elephants, a specific subset within the group Elephantidae, and located phylogenetically between the two living elephant groups (the African Loxodonta species and the Asian Elephas). This means that those behavioural and anatomical traits common to both living elephant groups should (in the absence of evidence to the contrary) be assumed present in mammoths. Mammoths were thus surely as complex and sophisticated in terms of social behaviour, noise-making, sensory abilities, intelligence and physiology as living elephants. I actually think it’s reasonable to speculate that – in ways – some mammoths were likely ‘more complex’ in terms of abilities and intelligence than living elephants in view of things like (probably) greater brain size and greater trunk complexity, plus they were clearly adapted for ecosystems and environments that were more complex, and more changeable, than the ones inhabited by elephants today.

Caption: some of the books you might like to consult if wanting to know more about Woolly mammoths. There are at least three editions of Adrian Lister and Paul Bahn’s Mammoths: Giants of the Ice Age; those shown here are those of 1995 and 2007. It’s my go-to work on this animal. Image: Darren Naish.
Finally… we think of the Woolly mammoth as a prehistoric animal. As a denizen of the Pleistocene (and one that seemingly originated between around 500,000 and 200,000 years ago, during the Chibanian or ‘Middle’ Pleistocene; read on), it of course is ‘prehistoric’ for a good chunk of its existence. But it also persisted beyond the Pleistocene and into the Holocene, famously surviving on Wrangel Island off Siberia until around 4000 years ago (Lister 1993, Vartanyan et al. 1993), and on St Paul Island off Alaska until about 8000 years ago (Guthrie 2004).
This makes the species a modern one that (in historic terms) only recently become extinct. It wasn’t just an ‘Ice Age’ animal, but one of the modern period – the Holocene – too. Incidentally, similarly young claims for Woolly mammoth persistence into the Holocene have more recently been made for mainland locations in Siberia and Canada. Argument remains over how reliable these dates are.

Caption: a key specimen in our understanding of the Woolly mammoth is the Beresovka mammoth, excavated in 1901 from the far north-east of Siberia and then transported across land all the way west to St Petersburg, a distance of over 10,000 km. As you can see, most of the soft tissues of the face were lost prior to excavation, but a large amount of soft tissue on the limbs and underside remained, as did various organs, a large amount of skin, and some hair. Image: public domain.

Caption: this is what the Beresokva mammoth display looks like (or… looked like, when this photo was taken) in St Petersburg’s zoological museum. I’ve never seen a detailed explanation of how much of this is real versus how much is a model, though at least one-third of the model’s exterior features the specimen’s original skin and hair (Lister & Bahn 2007, p. 52). The animal – an adult male in his 40s – seems to have broken its limbs when falling backwards into a crevasse and is positioned as if trying to climb back out. However, the body was moved by a landslide prior to discovery and this might also explain part of its posture and some of its damage. Image: Andrew Butko, CC BY-SA 3.0 (original here).
A quick look at Woolly mammoth evolution. Much of our knowledge of Woolly mammoth anatomy and evolutionary change comes from western Europe, and here the species is present in ‘final’ or ‘ultimate’ form from around 100,000 years until the end of the Pleistocene around 11,000 years ago (Lister 1996). Recent work has revealed the existence of individuals intermediate between the larger, taller Steppe mammoth M. trogontherii and the Woolly mammoth in north-east Siberia from around 700,000 years ago to about 200,000 years ago (Lister 2001, Lister et al. 2005, Lister & Bahn 2007). The main way of distinguishing Steppe and Woolly mammoths is in lamellar frequency, lamellae being the vertical plates in their molars. M. primigenius has more closely spaced, more numerous lamellae than M. trogontherii and other mammoths and seemingly evolved teeth of this sort due to the development of drier conditions in Siberia.

Caption: at left, an indication of how complex population structure seems to be across the Woolly mammoth, from Chang et al. (2017). Clearly, you can’t be expected to read the text on the cladogram here, but hopefully you can see the colours: the red samples are Asian Woolly mammoths; the green samples are North American ones; the blue samples are European ones (ignore the legend labelling blue as ‘M. jeffersoni’; I think that this is a typo for M. trogontherii). There’s a complex structure here, with multiple distinct geographically segregated clades but also multiple indications of introgression. At right, a very simple evolutionary tree, from Rohland et al. (2007), used to show how mammoths are very much part of the Loxodonta + Elephas clade, meaning that they are elephants.
While the picture is not yet entirely clear, it seems that mammoths with a reduced body size, higher lamellar count and increased hypsodonty (tooth height) – by definition, these are Woolly mammoths – had evolved from among M. trogontherii in Siberia by about 400,000 years ago. They then spread both west (to Europe) and east (to North America), ‘replacing’ older M. trogontherii populations in Eurasia by around 200,000 years ago (Lister et al. 2005, Lister & Shev 2015, Chang et al. 2017, Lister 2022). In some regions (like western Europe), this replacement looks rapid, but mixing and hybridization between the two might explain animals that look like ‘primitive Woolly mammoths’ or ‘advanced Steppe mammoths’. Incidentally, an independent movement into North America of Steppe mammoths likely explains the origin there of the Columbian mammoth M. columbi (Lister & Shev 2015), but that’s another story.

Caption: among the most important and interesting of permafrost Woolly mammoth specimens are the youngsters we’ve discovered. The stories behind how each met its end are very sad. At left is Dima, a male (6-12 months old) from the Magadan region of Siberia, discovered in 1977; at right, Lyuba, a female (just over 1 month old) from the Yamal Peninsula discovered in 2007. Images: public domain (original here); Ruth Hartnup, CC BY 2.0 (original here).
Out of America. Until recently, we’ve assumed that the Woolly mammoths present across Eurasia during the late Chibanian and Late Pleistocene were a continuous population. However, some genetic data indicates that something else happened. Debruyne et al. (2008) found that genetic haplotypes exclusive to North American Woolly mammoths replaced or superseded others across Eurasia from around 300,000 years ago onwards. They termed this the ‘Out of America’ hypothesis and argued that North American Woolly mammoths reinvaded Eurasia and thus restructured Woolly mammoth populations during the last part of their tenure. Chang et al. (2017) more recently published a massive genetic analysis of Woolly mammoths from many populations and found what looks like good support for Debruyne et al.’s model.
It shouldn’t be assumed that actual ‘replacement’ occurred here, as the North American invaders presumably hybridized with the older, Eurasian populations. I like imagining that these ‘American invaders’ had had time to evolve different looks and coat colours from Eurasian mammoths in which case they might have introduced new variants into the now messy Eurasian population… but I’m speculating.
What the Woolly mammoth was like. We know an unusual amount about the Woolly mammoth’s life appearance due to the finding of freeze-dried permafrost specimens from Siberia, from rock art created by our own species, and genetics, which has shed light on coat colour in life. A few basic traits of Woolly mammoth life appearance are obvious from its skeleton. It wasn’t especially big as proboscideans go, a large male being about 3.4 m tall at the shoulder, about 4.5 m long (without including the trunk) and probably between 4 and 6 tonnes in mass. The tusks – which were longer, more prominently curved and thicker in males – could be as much as 3.8 m long along the curve. Woolly mammoth tusks (and those of other mammoths too) curve outwards and downwards before sweeping sharply inwards and upwards.
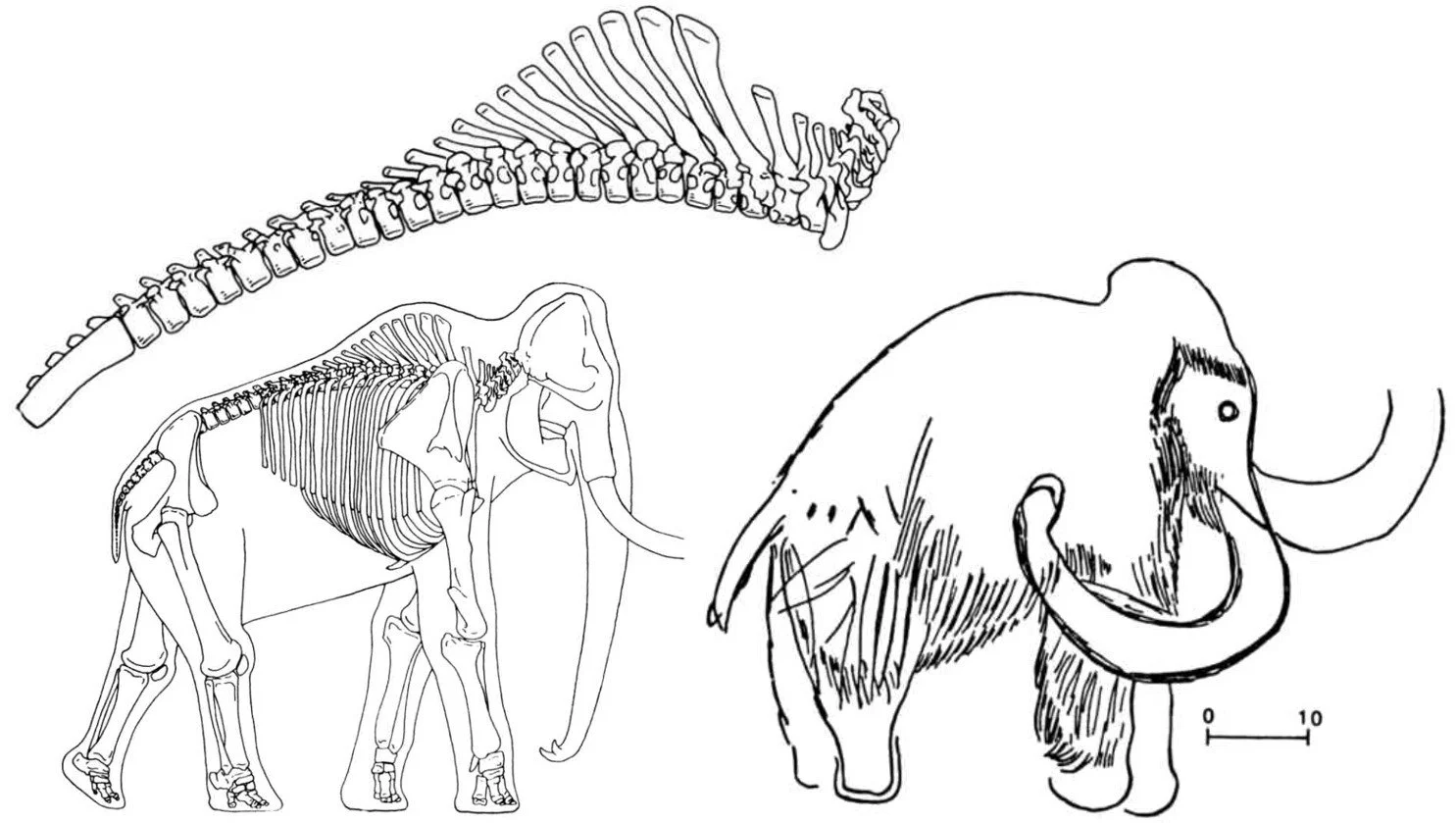
Caption: the Woolly mammoth has, or had, a very distinctive profile, the tall anterior thoracic neural spines being accentuated in height by a fatty hump, the skull being massively domed, and the body distinctly sloping from shoulders to hips. These features were very nicely illustrated by de Spiegeleire (1985). Those people who saw live Woolly mammoths noted these features too, and drew attention to them in the art they created. At right is a depiction of the famous Les Combarelles mammoth from Dordogne, France (from de Spiegeleire 1985). Other interpretations of this illustration show hanging hair beneath the chest and belly.
The profile of the Woolly mammoth was among the most distinctive of all mammoths. The skull was highly domed, with a single massively tall and rounded cranial prominence, and much shorter front to back than that of earlier mammoth species. A tall, fatty shoulder hump was prominent, and the body sloped sharply away from the hump, the hips being notably lower.
Both rock art and permafrost specimens reveal the presence of two ‘fingers’ at the trunk tip, this surely meaning that Woolly mammoths were especially good at picking up small objects like the herbs and grasses they fed upon. The dorsal ‘finger’ is longer (at 10 cm) and narrower than the ventral one (which is about 5 cm long) (Lister & Bahn 2007). An especially interesting and slightly controversial structure is a ‘mitten’ present about half-way along the trunk’s length at the sides of its ventral surface. It appears that two mobile lateral flaps allowed the trunk tip to be protected from cold when the trunk was partially rolled up. This structure is known from more than one permafrost specimen and does appear to have been a genuine piece of anatomy. Woolly mammoth ears were small, approximately similar in form to human ears, and mostly obscured by the surrounding long hairs.

Caption: several permafrost Woolly mammoth specimens show what the trunk’s end was like. At left is a diagrammatic representation of the Middle Kolyma mammoth, a specimen collected in 1924. A photo of the same specimen is shown in Lister & Bahn (2007, p. 91). At right, an effort to depict the trunk tip in life and showing the large size of the two ‘fingers’. Both images are from de Spiegeleire (1985).

Caption: the spectacularly preserved distal end of the trunk of the Maly Lyakhovsky Woolly mammoth from the New Siberian Islands, described by Grigoriev et al. (2017). This specimen is so fresh that its cut flesh is still red, as visible in B. The ‘fingers’ at the trunk tip are again well preserved. This specimen makes it look as if the trunk’s ventral surface, ventrolateral edges and tip were naked-skinned, rather than fully hair-covered. Maybe that was the case (maybe in some mammoths, at some times of year), but maybe this is due to damage and decomposition, since these permafrost specimens are never pristine. Images: Grigoriev et al. (2017).
Rock art and permafrost specimens show that Woolly mammoths had a coat formed of long guard hairs, the longest of which were around 40 cm along and would have hung from the animal’s upper surface and sides to create a shaggy ‘skirt’ superficially like that of musk ox and yak. Tighter, shorter hairs covered the limbs all the way down to the edges of the nails and foot pads. The tail was relatively short (at about 40 cm long), but long guard hairs (some as much as 60 cm long) grew from its edges and tip and it would usually have been mostly obscured within the pelage. This thick coat, combined with a fat layer 8-10 cm thick and genetics that indicate reduced sensitivity to cold (Lynch et al. 2015, Díez-del-Molino et al. 2023), mean that Woolly mammoths were well adapted to the extreme cold of Ice Age temperatures.
Today it’s well known that the reddish-brown coat colour conventionally applied to the Woolly mammoth might not be accurate but instead reflect degradation and colour change of preserved hairs over time. Genetics indicates that Woolly mammoths were mostly dark (exactly how dark is unsure though; it doesn’t mean black) and that lighter tones were possibly present in some individuals (Römpler et al. 2006, Workman et al. 2011), albeit very rare. Anatomical data from hairs renders things ambiguous though, since microscopic work still indicates a mostly pale colour to the outer hairs, in which case Woolly mammoths might have been straw-coloured, mid-brown or even mottled (Tridico et al. 2014).

Caption: the Woolly mammoth is a palaeo-superstar, and consequently a substantial number of models, figures and toys exist. Here are just a few of those from my own collection (TetZooTowers_collection on TikTok!). Image: Darren Naish.
As big, thick-coated animals that must have undergone seasonal changes in coat length and thickness across the course of the year, they were almost certainly similar to such modern species as Bactrian camels, musk ox and yak in that they would often have looked untidy, with large mats of shedding hair, tangled tufts at the edges of the coat, and filthy or stained areas around the genitals or back end. An interesting but undiscussed area concerns how greasy they were. They did have sebaceous glands – despite a long history of people denying this as a possibility – and were thus presumably good at repelling water. If you’ve interacted with a sufficient number of thick-coated big mammals, you’ll know that they can be surprisingly oily.

Caption: we might never know the precise details of their moulting schedule – if they had one – but it’s extremely likely that Woolly mammoths were sometimes untidy and with a decidedly complex look to their pelt, like these captive Bactrian camels (photographed at the Welsh Mountain Zoo). Images: Darren Naish.
Today, elephants are remarkable and magnificent animals that have a massive impact on their environment, are highly visible and audible due to their size and activities, and which surprise, enthral and impress us due to their social lives, intelligence and abilities. It’s hard, in some part of the world, to imagine that elephants much like the living ones, yet so different as well, lived in such places as England, France, Russia, or right across North America. Yet this was a normal aspect of what the world was like.
UPDATE: as if to emphasize the point that certain Steppe mammoths are hard to distinguish from certain Woolly mammoths, I am informed by Steve Zhang that the Ilford mammoth featured here is now regarded as probably belonging to the former species!
For previous Tet Zoo articles on proboscideans and on Pleistocene megafauna too, see…
How do you masturbate an elephant?, June 2008
The monster sheep that wasn’t, and other tales of African Bovini, May 2011
The Life Appearance of the Giant Deer Megaloceros, September 2018
Release the Fossil Pronghorns!!, November 2021
Armadillo Empire, Part 2: Fairies, Tolypeutines, and Where Glyptodonts Go, June 2025
Sloth World, 2025 (Part 1), October 2025
This blog benefits from your support. Thank you to those who help via patreon!
Refs - -
Debruyne, R., Chu, G., King, C. E., Bos, K., Kuch, M., Schwarz, C, Szpak, P., Gröcke, D. R., Matheus, P., Zazula, G., Guthrie, D., Froese, D., Buigues, B., de Marliave, C., Flemming, C., Poinar, D., Fisher, D., Southon, J., Tikhonov, A. N., MacPhee, R. D. E. & Poinar, H. N. 2008. Out of America: ancient DNA evidence for a New World origin of Late Quaternary Woolly mammoths. Current Biology 18, 1320-1326.
Grigoriev, S. E., Fisher, D. C., Obadă, T., Shirley, E. A., Rountrey, A. N., Savvinov, G. N., Garmaeva, D. K., Novgorodov, G. P., Cheprasov, M. Yu., Vasilev, S. E., Goncharov, A. E., Masharskiy, A., Egorova, V. E., Petrova, P. P., Egorova, E. E., Akhremenko, Y. A., van der Plicht, J., Galanin, A. A., Fedorov, S. E., Ivanov, E. V. & Tikhonov, A. N. 2017. A woolly mammoth (Mammuthus primigenius) carcass from Maly Lyakhovsky Island (New Siberian Islands, Russian Federation). Quaternary International 445, 89-103.
Guthrie, R. D. 2004. Radiocarbon evidence of mid-Holocene mammoths stranded on an Alaskan Bering Sea island. Nature 429, 746-749.
Lister, A. M. 1993. Mammoths in miniature. Nature 362, 288-289.
Lister, A. M.1996. Evolution and taxonomy of Eurasian mammoths. In Shoshani, J. & Tassy, P. (eds) The Proboscidea. Oxford University Press, Oxford, pp. 203-213.
Lister, A. M. 2001. “Gradual” evolution and molar scaling in the evolution of the mammoth. In The World of Elephants – International Congress, Rome 2001, pp. 648-651.
Lister, A. M. 2022. Mammoth evolution in the late Middle Pleistocene: The Mammuthus trogontherii-primigenius transition in Europe. Quaternary Science Reviews 294, 107693.
Lister, A. M. & Bahn, P. 2007. Mammoths: Giants of the Ice Age. Frances Lincoln Ltd, London.
Lister, A. M. & Sher, A. V. 2001. The origin and evolution of the Woolly mammoth. Science 294, 1094-1097.
Lister, A. M. & Sher, A. V. 2015. Evolution and dispersal of mammoths across the Northern Hemisphere. Science 350, 805-809.
Long, A., Sher, A., van Essen, H. & Wei, G. 2005. The pattern and process of mammoth evolution in Eurasia. Quaternary International 126-128, 49-64.
Römpler, H., Rohland, N., Lalueza-Fox, C., Willerslev, E., Kuznetsova, T., Rabeder, G., Bertranpetit, J., Schöneberg, T. & Hofreiter, M. 2006. Nuclear gene indicates coat-color polymorphism in mammoths. Science 313, 62.
de Spiegeleire, M. A. 1985. Figurations paléolithiques et réalité anatomique du mammouth (Mammuthus primigenius) essai d’interprétation. Bulletin de la Société royale Belge d’Anthropologie et de Préhistorie 96, 93-116.
Tridico, S. R., Rigby, P., Kirkbride, K. P., Haile, J. & Bunce, M. 2014. Megafaunal split ends: microscopical characterisation of hair structure and function in extinct woolly mammoth and woolly rhino. Quaternary Science Reviews 83, 68-75.
Vartanyan, S. L., Garutt, V. E. & Sher, A. V. 1993. Holocene dwarf mammoths from Wrangel Island in the Siberian Arctic. Nature 362, 337-340.
Workman, C., Dalen, L., Vartanyan, S., Shapiro, B., Kosintsev, P., Sher, A., Gotherstrom, A. & Barnes, I. 2011. Population-level genotyping of coat colour polymorphism in woolly mammoth (Mammuthus primigenius). Quaternary Science Reviews 30, 2304-2308.



