Yes … it’s time again to talk about the sloths, how I love them so. By the way, over recent months I’ve had to train myself to pronounce ‘sloth’ in the American way, not ‘sloathe’…

Caption: there are two very basic things to remember about sloths... that there are small, tree-climbing ones, and a substantial diversity of mostly larger or MUCH larger, extinct ones. Here are two exemplars of those facts: a captive Choloepus at left, and a museum skeleton (a cast) of the South American giant Megatherium (at the Natural History Museum, London) at right. Images: Darren Naish.
Following the armadillo-themed articles that appeared here within recent months, it only seems fair to give sloths some coverage too. And such a thing is well overdue, since I haven’t written properly about sloths for years now. Indeed, a sloth-themed review article I published back in 2005 (Naish 2005) is now chronically out of date with respect to what it says about sloth phylogeny, diversity and history.
Sloths are a big group and there’s a lot to say about them, so my aim here is to focus on a few select highlights. I can say now that – yes – sloths are a group I’ve been considering at length within recent years thanks to Prehistoric Planet Ice Age, due out in late November 2025. More on that in due time.

Caption: there’s a fair bit about sloths in the Tet Zoo archives, but it’s now hard to find and much of it has been ruined by removal of images and such. Here are two of the relevant articles, one from 2007, one from 2010. Of relevance here is that I’m gradually building a new archive of Tet Zoo articles (it’s here), aiming where possible to find intact versions at the internet archive.
Sloths of the Caribbean. We’ve known since the 1860s that sloths inhabited various of the Caribbean islands, since that’s when Megalocnus rodens – initially deemed notable because of its rodent-like anterior dentition – was described from the Pleistocene of Cuba. Megalocnus was similar in size to an average bear and seems from limb proportions and foot shape to have been a terrestrial, ground-feeding animal.
Later finds showed that similar, related sloths occurred in Puerto Rico (Acratocnus) and Hispaniola (other species of Acratocnus, plus Parocnus and Neocnus); additional taxa lived in Cuba too (Imagocnus, and other species of Acratocnus and Neocnus). As many as six species lived on certain of these islands (Viñola-Lopez et al. 2021). These were generally smaller than Megalocnus (the Neocnus species were tiny; smaller than most living tree sloths at less than 15 kg) and some (like Acratocnus) were especially short-snouted. Excepting Imagocnus (which is Miocene), these are animals of the Late Pleistocene and Holocene, and remains of Megalocnus and Parocnus (plus of indeterminate species) show that some were alive 6000-5000 years ago (Steadman et al. 2005). They’re modern and recently extinct, not prehistoric.

Caption: megalocnoid skulls from Hispaniola as illustrated by MacPhee et al. (2000). (A) Acratocnus ye, (B) Neocnus dousman and (C) N. toupiti. Note the scale bars: the skulls of these animals are (relatively!) tiny (it’s 9.7 cm in N. toupiti). The skulls of these animals are also narrow across the snout. Image: MacPhee et al. (2000).
All the sloths mentioned here share traits and have traditionally been grouped together in Megalocnidae. A popular idea on their phylogenetic placement has been that they’re part of Megalonychidae – the group anchored on Megalonyx, a particular famous North American ground sloth – and, within that group, close to Choloepus, the extant two-fingered* sloths (e.g., Gaudin 1995, 2004, Pujos et al. 2007). This seems logical in view of superficial similarities in skull form and the small size and probable semi-arboreal lifestyle of Caribbean taxa like Neocnus.
* I’ve always disliked the terms ‘two-toed’ and ‘three-toed’ for extant sloth species, given that we’re talking about fingers, not toes… so I welcome new efforts to change the common names of these animals.
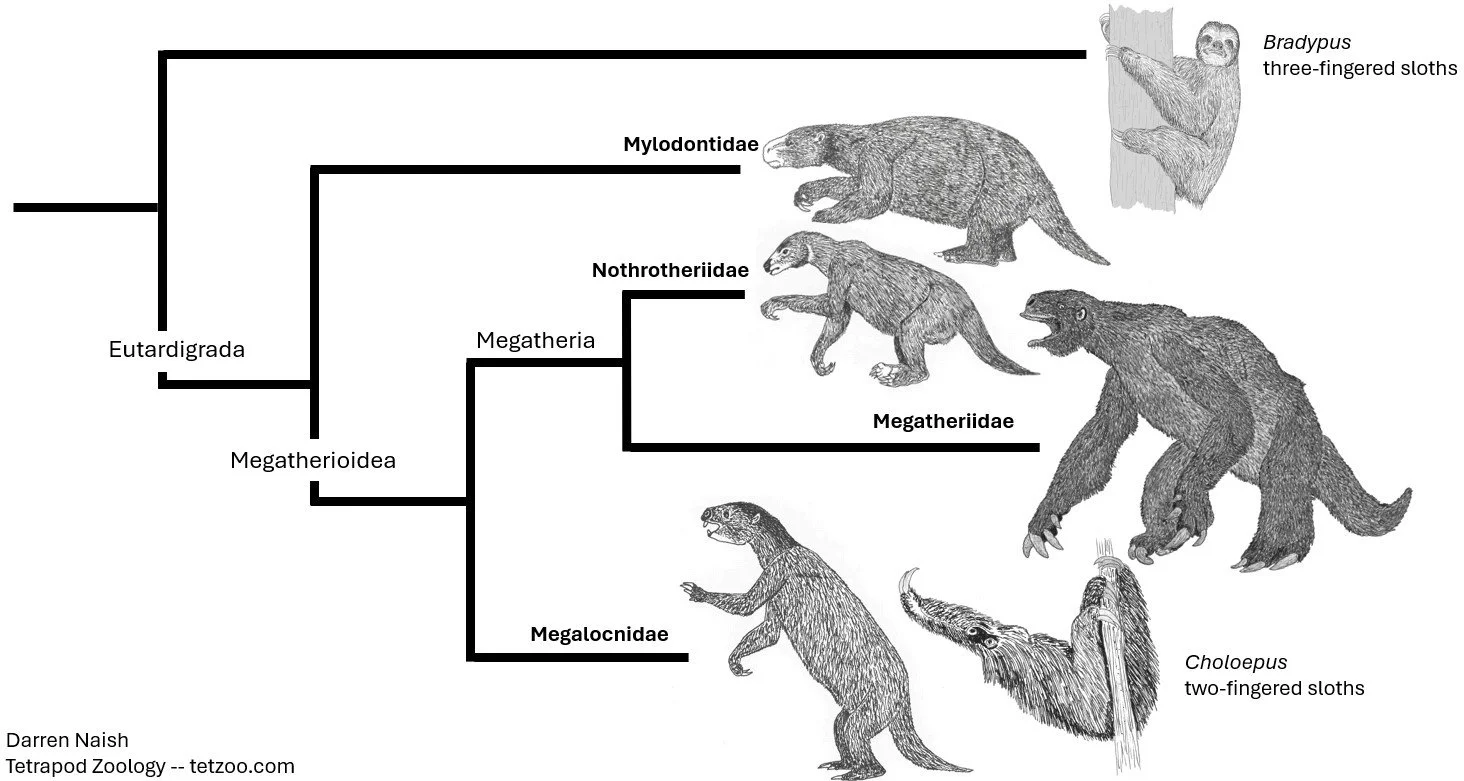
Caption: a simplified version of the morphology-based view of sloth phylogeny generally thought correct prior to the 2010s. Bradypus was regarded as the earliest diverging sloth lineage (in which case it has a suspiciously long ghost lineage), nothrotheres and megatheriids were considered close kin, and Choloepus was a megalocnid and hence part of one of the youngest sloth groups. Images: Darren Naish.
However, molecular results indicate that this is completely wrong. Instead, it seems that the Caribbean sloths represent an entirely distinct lineage that diverged early on within sloth history, way back in the Eocene (Delsuc et al. 2019, Presslee et al. 2019, Viñola-Lopez et al. 2021). They’re thus not close to Megalonyx or kin, nor to Choloepus, and their early colonization of the Caribbean has required a rethink on how they became distributed. In addition, an Oligocene divergence of the Aractocnus and Parocnus lineages (it seemingly occurred around 30 million years ago) has led to a revised taxonomy where Acratocnidae and Parocnidae are regarded as the main constituent groups within an ‘up-ranked’ Megalocnoidea (Delsuc et al. 2019).
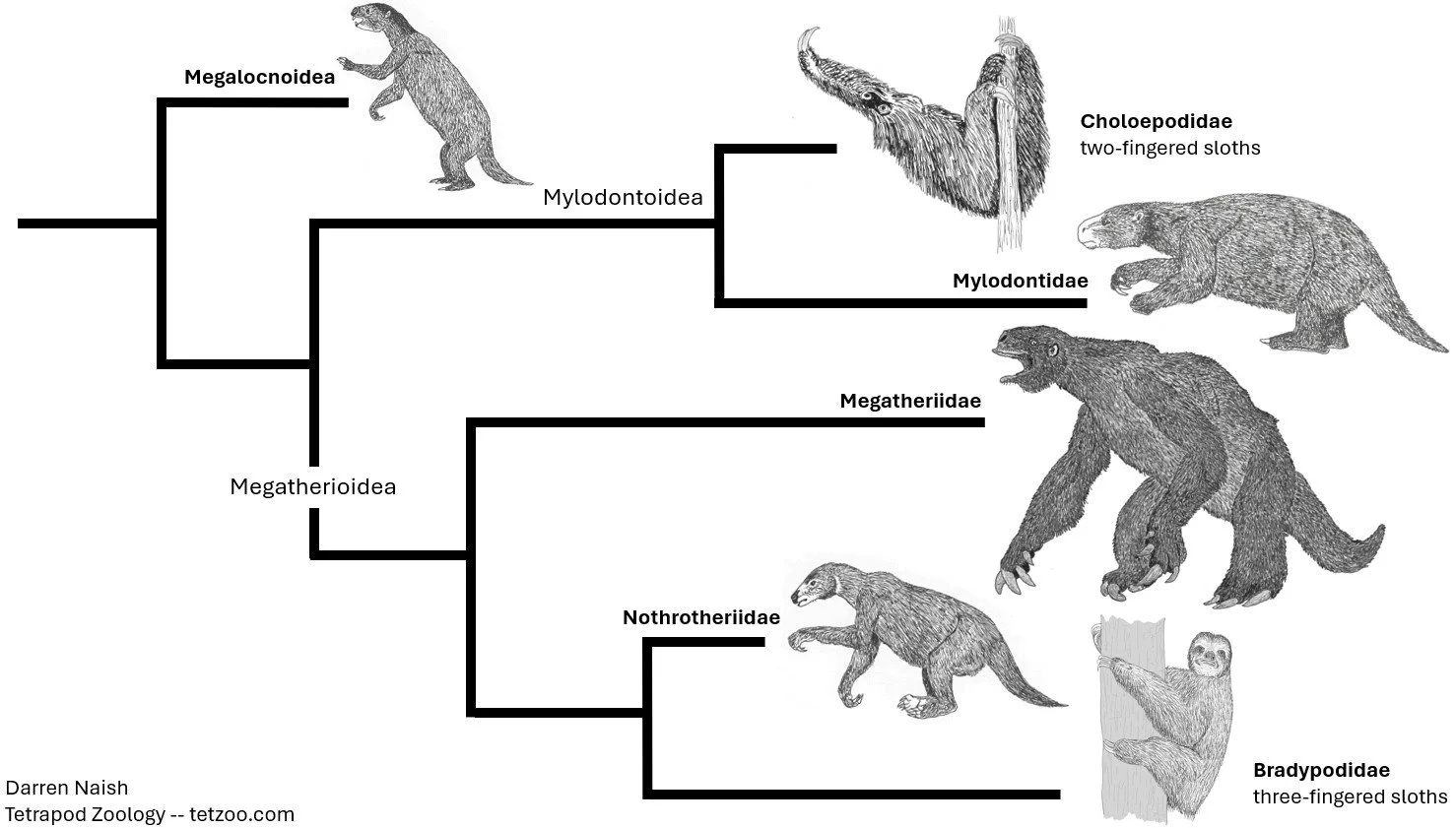
Caption: molecular data indicates that the pattern of sloth evolution was rather different from what we thought based on anatomy. Caribbean sloths are the sister-group to the rest of the clade and Choloepus is not close to this group (instead being closer to mylodontids); Bradypus does not belong to an especially archaic lineage but is instead within the clade that includes megatheres and nothrotheres. Image: Darren Naish.
Big megatheres: furry, naked or both or neither? Within recent years, the view that extinct sloths of (essentially) all sorts should be imagined as shaggily-pelted has come into question. We have direct evidence that at least some mid-sized, extinct sloths (namely Mylodon and Nothrotheriops) were like this, since of course we have preserved segments of their pelt. But was it true for all of them, in particular the very big ones that lived (in parts of their ranges) in tropical climes?

Caption: one of the famous fragments of Mylodon skin (with pelt) from Cueva del Milodón Natural Monument in Chilean Patagonia, specifically the one collected by Francisco P. Moreno in 1897. Despite claims that sloth fur is coarse and stiff, you’ll note that the hair here actually has a soft and lustrous look. Small bony nodules are embedded on the skin’s inner surface. This specimen is in the collections of the Natural History Museum, London. Some dung from the same location is visible at back. Image: Darren Naish.
On a few occasions, experts have argued that giant taxa – in particular the megatheres Megatherium and its close relative Eremotherium – were more sparsely haired, perhaps recalling extant elephants or domestic pigs in hair coverage. Supposedly, they would have benefited from this, since a thick pelt would result in heat stress (Fariña 2002). However, sloths have a relatively low metabolism compared to elephants and pigs (and this is apparently true even if we accept arguments about extinct sloths having higher metabolisms than conventionally assumed; Varela et al. 2024), so… are we sure that it’s ‘one integumentary rule applies to all’ when it comes to physiological arguments of this sort?
Exactly this was examined by Deak et al. (2025). By simulating the climatic regimes that would have been experienced by megatheres when they were alive and combining this with a metabolic rate, Deak et al. (2025) found that megatheres would have suffered from cold stress across large swathes of their range if they had sparse or absent fur. A dense, long coat would likely have caused heat stress in tropical places, but a dense and thick pelage was otherwise needed even for these giants. In view of this, Megatherium and Eremotherium might even have been variable in integument across their ranges (Deak et al. 2025).

Caption: at left, a graph from Deak et al. (2025) showing the temperature regimes experienced by select extinct sloths. A famous ‘tropical’ sloth like the gigantic Eremotherium experienced warm temperatures in part of its range, but it lived in places that were cool and cold as well. At right, Eremotherium laurillardi skeleton at Houston Museum of Nature and Science. Images: Deak et al. (2025); Kamraman, CC BY-SA 2.5 (original here).
Marine sloths. If you’re at all similar in age to myself, you’ll remember how exciting it was to see the 1990s discovery of Thalassocnus natans, a coastal South American sloth, initially described from the Upper Miocene of Peru, that possesses adaptations throughout the skeleton for swimming (de Muizon & McDonald 1995). Additional, geologically younger Thalassocnus species were discovered later on, some of which look to be more specialized for swimming than T. natans (de Muizon et al. 2003, 2004a).

Caption: the 1995 publication of Thalassocnus natans from Peru caused quite the sensation. As you see from this articulated skeleton (on show at Muséum National d'Histoire Naturelle, Paris), it’s not tremendously modified relative to other sloths: adaptations in the tail, hindlimb and skull are consistent with a wading, aquatic feeding lifestyle. However, the discovery since 1995 of other thalassocnine species show that members of this lineage became increasingly modified for marine life over time. Image: FunkMonk, CC BY-SA 3.0 (original here).
All species are mid-sized as sloths go, with body length estimated at being between 2.5 and 3 m long. In the youngest species – T. yaucensis from the Late Pliocene – the bones at the front of the skull are elongate and narrow and indicative of flexible lips specialized for marine feeding. T. yaucensis also lacks microwear indicative of sand ingestion and hence was apparently feeding in deeper water than the less specialized species (de Muizon et al. 2004b). In the rest of the skeleton, the radius superficially recalls that of pinnipeds (de Muizon et al. 2003). As tempting as it is to imagine these last members of the group as pelagic ‘seal-sloths’, if you like, it might be that they were more adapted for shallow-water floating and walking and not under evolutionary pressure to evolve a fusiform shape. They’re not unique to Peru by the way, later finds demonstrating a presence in Argentina and Chile.
This lineage – grouped together as Thalassocninae – was originally included within the nothrotheres (read on for more on that group), and some studies do still support this view (Pujos et al. 2016). However, other studies place them within Megatheriidae (Amson et al. 2017), the sloth group best known for including the giant megatheres discussed above. Incidentally, suggestions have been made that other extinct sloths were adapted for life in water. Lestodon – a robust-skulled mylodontid with thick and prominent caniniform teeth – has been compared to hippos on occasion and hence imagined as an amphibious animal. I’d like to see this formally tested though.

Caption: Lestodon of the Pliocene, Pleistocene and Holocene of Brazil, Paraguay, Uruguay and Argentina is a large and hilariously chunky mylodontid sloth with a notably broad, robust rostrum. The suggestion has been made several times that it might have been hippo-like in some aspects of behaviour and ecology. Images: skeletal reconstruction modified from Tomassini et al. (2020); the skull illustration is from a paper by Greg McDonald but I’m unsure of its original provenance, CC BY 4.0 (original here).
Shasta ground sloths and other nothrotheres. Excluding the thalassocnines, nothrotheres – or nothrotheriids to be more formal – are a group of small- and mid-sized, mostly terrestrial sloths that lack the specializations typical of other groups and share relatively narrow cheek teeth, a prominent and asymmetrical vomerine keel on the bony surface of the pharyngeal cavity (Gaudin 2004), bulbous mastoid processes at the back of the skull and a list of other traits in the skull, forelimb and ankle. They have slender, more elongate arm bones than most other extinct sloth groups. Genetic data shows that they aren’t archaic sloths, but a relatively young group akin to megalonychids and the extant three-fingered Bradypus (Delsuc et al. 2019).

Caption: skeleton of the familiar nothrothere Nothrotheriops, aka Shasta ground sloth, as displayed at Peabody Museum. At right, a 1938 photo of Nothrotheriops dung from Rampart Cave in Arizona. This dried dung is simply old and dry and has never been buried, so I don’t think people have ever referred to it as ‘fossil’. Images: public domain (originals here and here).
The history of thoughts on nothrothere taxonomy and phylogeny is complex and involves more taxa than I have time to talk about here. A number of Miocene taxa appear to be early-diverging members of the group outside a Pliocene-Holocene clade (Nothrotheriinae) that includes Nothrotherium itself, a well-studied animal from the Pleistocene and Holocene of South America. Coprolites and isotopic data show that this sloth inhabited parkland-like forests and ate dry-adapted fruits and leaves. Similar things can be said for the related Nothrotheriops, one species of which (N. shastensis) is known as the Shasta ground sloth following its early 20th century discovery in Shasta County, California. Copious air-dried dung belonging to N. shastensis is known from the American south-west and shows that these animals used caves and rocky overhangs as refuge or resting sites. Related nothrotheres include Aymaratherium from the Pliocene of Bolivia, Pronothrotherium from the Miocene and Pliocene of Brazil and Mionothropus from the Miocene of Peru (De Iuliis et al. 2011, Pujos et al. 2016).

Caption: cladogram and map showing nothrothere evolution, from Pujos et al. (2016). This work shows nothrotheres as a mostly Miocene radiation of non-equatorial South America, one lineage of which migrated into North America. It remains controversial whether thalassocnines are nothrotheres, since they also share traits with megatheres. Image: Pujos et al. (2016).
That was a very brief look at just a few aspects of extinct sloth diversity and biology. There is so much more to say and I absolutely must come back to them in time. Watch this space.
For previous Tet Zoo articles on xenarthrans, see…
Five things you didn’t know about armadillos, June 2007
What was that skull? (on glyptodonts), May 2008
I, Priodontes, the tatuasu, September 2008
Predation and Corpse-Eating in Armadillos, September 2010
The Fate of the Woolly Long-Nosed Armadillo of Peru, April 2024
Of Zaedyus, the Pichi, May 2025
Armadillo Empire, Part 1: of Euphractines and Eutatines, June 2025
Armadillo Empire, Part 2: Fairies, Tolypeutines, and Where Glyptodonts Go, July 2025
Refs - -
Amson, E., Muizon, C. de & Gaudin, T. J. 2017. A reappraisal of the phylogeny of the Megatheria (Mammalia: Tardigrada), with an emphasis on the relationships of the Thalassocninae, the marine sloths. Zoological Journal of the Linnean Society 179, 217-236.
De Iuliis, G., Gaudin, T. J. & Vicars, M. J. 2011. A new genus and species of nothrotheriid sloth (Xenarthra, Tardigrada, Nothrotheriidae) from the Late Miocene (Huayquerian) of Peru. Palaeontology 54, 171-205.
Fariña, R. A. 2002 Megatherium, the hairless: appearance of the great Quaternary sloths (Mammalia; Xenarthra). Ameghiniana 39, 241-244.
Gaudin, T. J. 1995. The ear region of edentates and the phylogeny of the Tardigrada (Mammalia, Xenarthra). Journal of Vertebrate Paleontology 15, 672-705.
Gaudin, T. J. 2004. Phylogenetic relationships among sloths (Mammalia, Xenarthra, Tardigrada): the craniodental evidence. Zoological Journal of the Linnean Society 140, 255-305.
Muizon, C. de & McDonald, H. G. 1995. An aquatic sloth from the Pliocene of Peru. Nature 375, 224-227.
Muizon, C. de, McDonald, H. G., Salas, R. & Urbina, M. 2003. A new early species of the aquatic sloth Thalassocnus (Mammalia, Xenarthra) from the Late Miocene of Peru. Journal of Vertebrate Paleontology 23, 886-894.
Muizon, C. de, McDonald, H. G., Salas, R. & Urbina, M. 2004a. The youngest species of the aquatic sloth Thalassocnus and a reassessment of the relationships of the nothrothere sloths (Mammalia: Xenarthra). Journal of Vertebrate Paleontology 24, 387-397.
Muizon, C. de, McDonald, H. G., Salas, R. & Urbina, M. 2004b. The evolution of feeding adaptations of the aquatic sloth Thalassocnus. Journal of Vertebrate Paleontology 24, 398-410.
Naish, D. 2005. Fossils explained 51: sloths. Geology Today 21 (6), 232-238.
Presslee, P., Slater, G. J., Pujos, F., Forasiepi, A. M., Fischer, R., Molloy, K., Mackie, M., Olsen, J. V., Kramarz, A., Taglioretti, M., Scaglia, F., Lezcano, M., Southon, J., Feranec, R., Bloch, J., Hajduk, A., Martin, F. M., Gismondi, R. S., Reguero, M., Muizon, C. de, Greenwood, A., Chait, B. T., Penkman, K. & MacPhee, R. D. E. 2019. Palaeoproteomics resolves sloth relationships. Nature Ecology & Evolution 3, 1121-1130.
Pujos, F., de Iuliis, G., Argot, C. & Lars, W. 2007. A peculiar climbing Megalonychidae from the Pleistocene of Peru and its implication for sloth history. Zoological Journal of the Linnean Society 149, 179-235.
Steadman, D. W., Martin, P. S., MacPhee, R. D. E., Jull, A. J. T., McDonald, H. G., Woods, C. A., Iturralde-Vinent, M. & Hodgins, G. W. L. 2005. Asynchronous extinction of late Quaternary sloths on continents and islands. Proceedings of the National Academic of Sciences 102, 11763-11768.
Viñola-Lopez, L. W., Suárez, E. E. C., Vélez-Juarbe, J., Milan, J. N. A. & Bloch, J. I. 2021. The oldest known record of a ground sloth (Mammalia, Xenarthra, Folivora) from Hispaniola: evolutionary and paleobiogeographical implications. Journal of Paleontology 96, 684-691.



