Those of you specially interested in the behaviour and biology of Mesozoic dinosaurs will be well aware of the dinosaur cognition debate that’s been going on in the animal cognition literature…

Caption: a wholly speculative effort to portray potential cognition in an animal like a non-bird theropod. Which of these dotted lines is more realistic… or, is it totally misleading, perhaps wrong, to imagine or portray things this way? Images: Darren Naish.
As discussed at Tet Zoo back in 2024 (go here), things started when neuroscientist Suzana Herculano-Houzel argued that such extinct dinosaurs as Tyrannosaurus might have had primate-like numbers of neurons, and thus might have been similar to anthropoids in behavioural traits and overall intelligence (Herculano-Houzel 2023). I don’t need to say that these conclusions were surprising, the consensus view among those working on extinct dinosaurs otherwise thinking they were potentially similar in intelligence to extant squamates, crocs, and perhaps to such birds as ratites and gallinaceous birds.
I’m part of a team that pushed back on the details that Herculano-Houzel used to build her case: Kai Caspar, Cristian Gutiérrez-Ibáñez, Grant Hurlburt, myself and others showed that Herculano-Houzel’s brain size estimates were likely incorrect, that her neuron count estimates were unreliable and based on erroneous assumptions, and that the neuron counts she estimated weren’t clearly linked to anthropoid-like cognitive abilities anyway (Caspar et al. 2024). We published in The Anatomical Record and our article is open access. Concepts like ‘intelligence’ and ‘overall cognitive complexity’ are still difficult to quantify and describe in any meaningful way, so our takehome as expressed in popular sources (like this blog) was that animals like tyrannosaurs were more likely ‘reptile smart’ than ‘primate smart’… It is important to note, however, that it’s very hard to find us stating exactly this in our article. Consider the following statements from Caspar et al. (2024)…
“Cognitive traits identified exclusively in birds or crocodiles cannot simply be extrapolated to Mesozoic dinosaurs with any degree of certainty since they might represent crown group apomorphies. Although it might be appealing to hypothesize that cognitive patterns found among modern palaeognaths are representative for their maniraptoriform forerunners … this idea is (in most cases) not testable and should hence not be disseminated uncritically.”
“…. the extant archosaurian groups leave us in a rather suboptimal position to infer cognitive traits in non-avian dinosaurs.”
Our article (Caspar et al. 2024) was, of course, far from the end of the discussion. It turns out that people working elsewhere in animal cognition have also been following the argument, and – yes – they have thoughts.

Caption: ongoing studies of extant non-bird reptiles show that there’s a lot going on here in cognitive terms, and at least some are explorative, investigative creatures that are good at learning. It’s increasingly well known that monitor lizards (like the Komodo dragon at left) and iguanians like bearded dragons (like the pet one at right) pass numerous tests relating to 'intelligence’. Images: Darren Naish.
Jensen et al. (2025) respond. Thomas Rejsenhus Jensen of Lund University’s Cognitive Zoology Group, plus colleagues, responded; while not agreeing with Herculano-Houzel (2023) outright, part of their argument was that Caspar et al. (2024) might be criticized for creating the impression that a typological view holds sway in animal cognition; that reptiles (like non-bird dinosaurs) can only be ‘reptile-like’, for example. Instead, they point out, cognition involves multiple shades of grey in addition to a plasticity that relates to the specific lifestyle and adaptations of a species (Jensen et al. 2025)… a position, by the way, that we agree with. T. rex should not, then, be considered either ‘baboon-like’ or ‘crocodile-like’ but ‘T. rex-like’ and it’s misleading to use such terms as “smartness” “intelligence”, “baboon-like cognition” and “crocodile-like cognition” (Jensen et al. 2025).

Caption: “these animals are like x”, “no, they’re like y”, “no, they’re like what they’re like”. The articles shown here are from a debate that occurred in the journal Evolution between 1970 and 1974.
In addition, they argued that we’d ignored the ‘endothermic brain hypothesis’ and also suggested the presence of a ‘cognitive arms race’ as a potential explanation for the evolution of convergent neurocognitive traits in mammal-line and bird-line animals in the Mesozoic (Jensen et al. 2025).
There’s a lot to consider here, and late last year another article was published on this topic, this one being a response to Jensen et al. (2025) from Kai Caspar, Cristian Gutiérrez-Ibáñez, Grant Hurlburt, myself and others (Caspar et al. 2025). We took issue with several of Jensen et al. (2025)’s key points. Here, I summarise these and explain how we responded. Our article was again published in The Anatomical Record and is again open access. We owe substantial thanks to the editorial team at The Anatomical Record for swift and efficient handling, plus to our reviewers and colleagues.

Caption: the topic being covered here is of broad interest and will get covered in the popular and semi-technical press. In view of this, what language do we use and which comparisons do we make when discussing the subject? The articles here are from CBBC and Tet Zoo; I can no longer find ‘Scientists change their minds about Tyrannosaurus rex yet again’ (a similar article by the same author is here).
On typology and non-bird dinosaurs as ‘reptile-brained’. As noted above, Jensen et al. (2025) accused us (Caspar et al. 2024) of simplifying or underselling the issue of dinosaur cognition. But this is complicated by the fact that the discussion as a whole is occurring in two locations: (1) the technical literature, and (2) the popular sphere (blog articles like the one you’re reading now, and in the popular press, since journalists enjoy covering this topic). Nowhere in our technical article did we (Caspar et al. 2024) use the terms that might be associated with a typological view: yes, we drew attention to the fact that extinct dinosaurs are often more ‘crocodylian-like’ in brain form and proportional size than birds, but we stated a nuanced position when it comes to what this might mean for cognition, as is clear from the quotes from Caspar et al. (2024) provided above.
When it comes to summarising the situation in popular sources, I don’t see an issue with using terms like ‘smartness’ and ‘intelligence’ so long as caveats, or quote marks (as here), are used. It remains difficult, if not impossible, to quantify or precisely measure overall cognitive complexity, meaning that approximate comparisons to living taxa are about the best way we have of describing things. Saying that dinosaurs like tyrannosaurs might have been approximately similar in intelligence to certain lizards or crocodylians, for example, is reasonable based on what we know (and, as I’ve emphasized here, not ‘a bad thing’ in terms of what it might mean for behavioural complexity, adaptability and so on). Jensen et al. (2025) appeared under the impression that the notion of ‘reptile-brained’ (non-bird) dinosaurs had been abandoned by scientists working on these animals, but that’s not so, as we demonstrated via quotes and citations from recent work on dinosaur biology (Caspar et al. 2025).

Caption: there’s a popular idea that palaeognaths (which include ratites, like this rhea) are not real birds and don’t do birdy things. Having spent a lot of time with them (cassowaries especially)… no. This captive rhea spent a good while engaging in what sure looked like object play with a feather. An argument has been made that some extinct dinosaurs were similar to ratites in cognitive terms. Images: Darren Naish.
An aside on nomenclature. As someone who sometimes publishes on the fossil record and diversity of crocodile-line archosaurs, I always make a point of using the specific terms that have been established in the technical phylogenetic literature. The term crocodile applies to one specific group of these animals and people – and this applies especially to palaeontologists – really, really need to wean themselves off the childish idea that it can be used indiscriminately.
The crown-group – the one that includes alligators, crocodiles and their close relatives – is Crocodylia, which we use to avoid confusion with the old ‘Crocodilia’, a term used in the past for the entire group (meaning Crocodylomorpha, and incorporating a massive diversity of extinct archosaurs that are wholly different in shape and behaviour from crocodylians). In short, modern crocodiles, alligators are kin should be referred to collectively as crocodylians. Back to the main issue at hand…
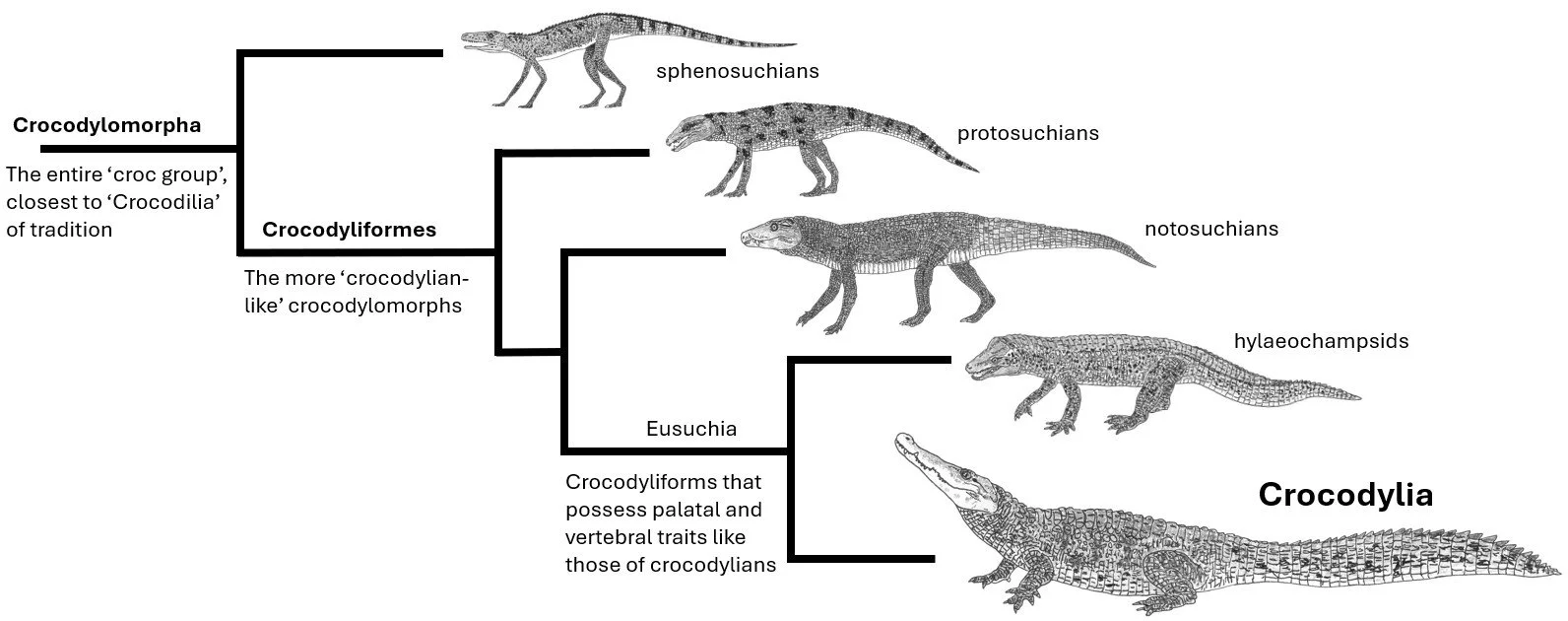
Caption: a substantially simplified cladogram showing relationships within crocodylomorphs. There is still a tendency within the natural history and zoological literature for the crown-group to be termed the ‘crocodilians’ but the name ‘crocodylian’ is more firmly established within phylogenetic literature. The incorrect tradition of referring to any and all of these animals as ‘crocodiles’ also needs to be put to bed. Crocodiles are a specific group of crocodylians. Image: Darren Naish.
Is the ‘endothermic brain hypothesis’ valid for Mesozoic vertebrates? The main reason we hadn’t mentioned the endothermic brain hypothesis or EBH in our initial article (Caspar et al. 2024) is that it hadn’t been proposed in print when we were writing. Its first published outing was in Osvath et al. (2024), which appeared in November 2024: Caspar et al. (2024) appeared in April 2024. Simply put, the EBH argues that whole-body tachymetabolic endotherms have evolved unusual cognitive traits (like larger brain size and higher neuron counts) in step with their need to find greater amounts of fuel for their elevated metabolisms, and that they operate via an advanced ‘model-based’ system of optimized foraging that requires greater cognitive processing than that present in other animals (Osvath et al. 2024).
Our main point of response (Caspar et al. 2025) is that robust evidence for a link between the evolution of endothermy and a marked change in brain anatomy (especially one measurable in fossils) is lacking. Fossils simply don’t show an anatomical ‘upgrading’ in brain form or complexity occurring in step with the hypothesised development of endothermy among the relevant groups. In addition, the idea that a modified (as in, proportionally big or especially neuron-dense) brain is associated with endothermy is countered by fossil pseudosuchians (which apparently combined endothermy with proportionally small brains) and big-brained, neuronally dense but ectothermic fishes (Caspar et al. 2025).

Captions: these images are here to remind you that there are actinopterygians (ray-finned fishes) and chondrichthyans (cartilaginous fishes) with proportionally enormous brains and extremely high neuron densities. How do these fit into patterns proposed for terrestrial animals like archosaurs? Images: opencage, CC BY-SA 2.5 (original here); Shiyam ElkCloner, CC BY-SA 3.0 (original here).
Was there a Mesozoic ‘cognitive arms race’? Jensen et al. (2025) proposed that the evolution of endothermy in synapsids and archosaurs might have been engaged in a “cognitive competition for food” and that this “could have boosted success compared to less cognitively apt competitors”. They were inspired by Benton’s (2021) very useful overview on the evidence for endothermy in Permo-Triassic synapsids and archosaurs, in which a ‘Triassic arms race’ relating to postural and locomotor changes was suggested.
Caption: mammal-line and bird-line animals interacted continuously throughout the Mesozoic, as they do today. But is there any evidence that anything like a cognitive ‘arms race’ was underway? This image, from the Apple TV series Prehistoric Planet, shows an interaction between a troodontid theropod and a multituberculate. Image: © Apple / BBC Studios.
There may or may not be merit to this idea. But even if there is, the fossil record does not show a concomitant cognitive ‘up-grade’ occurring in either group at the right time. “The proposal is thus implausible to us” (Caspar et al. 2025). See the article for the full discussion and relevant data.
Neuron counts and relative brain size again. Finally, we end our article by discussing the fact that neuron counts are too ‘noisy’ a metric to be especially reliable when it comes to making inferences about cognition. Jensen et al. (2025) criticized us for apparently questioning the idea that neuron counts are linked to cognitive ability, but we didn’t exactly do that: our point instead was that neuron counts are one among many things that need to be considered, and that they can’t be used alone as a proxy in making conclusions (as they were by Herculano-Houzel) (Caspar et al. 2024, 2025).
In fact, despite the argument from Jensen et al. (2025) that a clear link between raw neuron count and cognition exists, it’s still difficult to pin down the tightness of this correlation and determine whether absolute or relative neuron count is more important. For birds, most work indicates that relative brain size holds sway. Big members of animal groups have high neuron counts relative to small members of those groups because high counts correlate with overall size and don’t clearly grant cognitive advantages (Caspar et al. 2025).
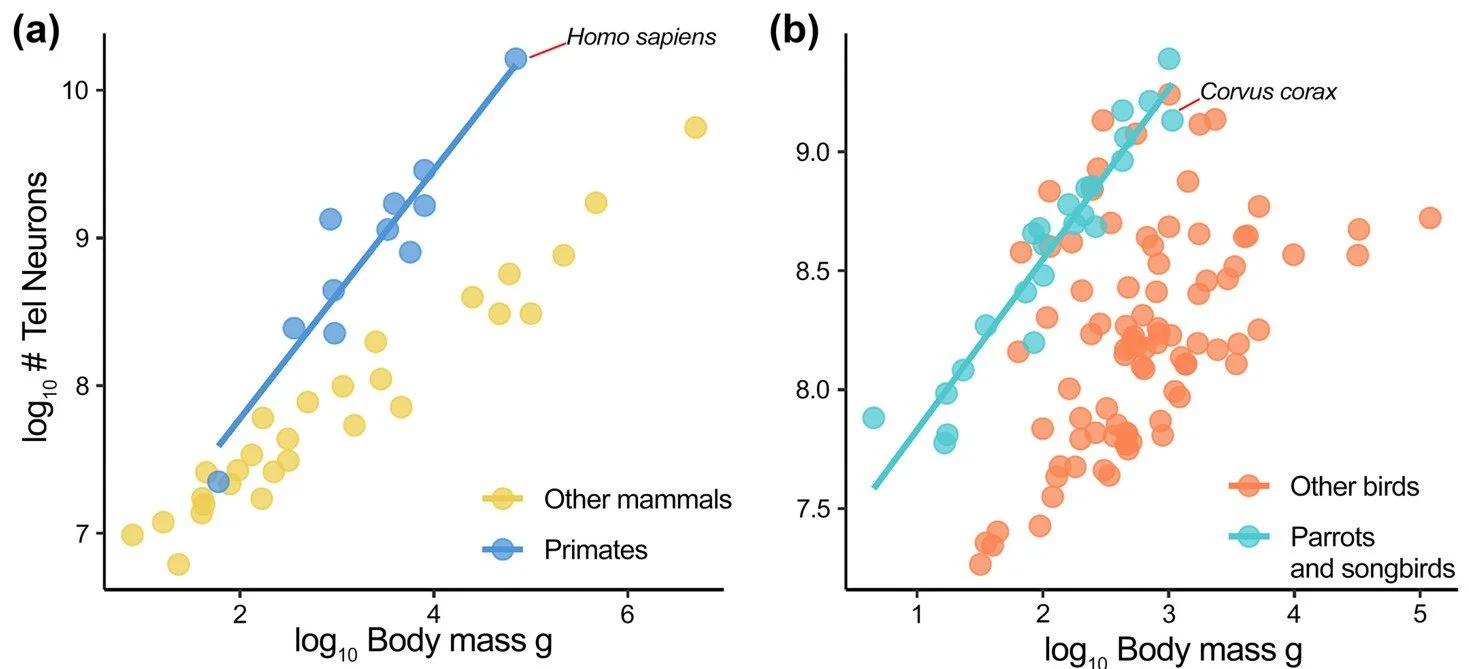
Caption: graphs from Caspar et al. (2025) showing neuron count in the telencephalon plotted against body mass in mammals (at left) and birds (at right). The line in the mammal graph shows the regression line for primates, showing that Homo sapiens has a neuron count expected for its body mass. The line in the bird graph shows likewise for Psittacopasserae, here showing that Corvus corax demonstrates the same thing within its clade. Image: Caspar et al. (2025).
Clearly, much remains to be learnt about the anatomy of dinosaur brains and what that anatomy does, or does not, mean for behaviour and cognition. The notion that we can make simple inferences about cognition based on overall physiology (the ‘endothermic brain hypothesis’) or that brain enlargement and hence behavioural complexity occurred as the consequence of a Mesozoic ‘arms race’ are both, however, so flawed that they should probably be considered wrong, and sections of our new article are devoted to discussing and refuting these ideas (Caspar et al. 2025).
As should be clear by now, this is very much an active area of debate and consideration. It’s difficult not to be intrigued and even excited by claims that Mesozoic dinosaurs possessed “largely unknown neurocognitive functions approaching those seen in birds” (Jensen et al. 2025). But on dinosaur cognition overall, these animals quite probably overlapped with turtles, squamates and crocodylians as well as birds (Caspar et al. 2024, 2025), the great caveat being that our understanding of cognitive traits in so many of these animals remain very much rudimentary. Again, this mustn’t be interpreted to mean that the extinct animals were deficient or poor in performance given what we currently think about learning, complexity and memory in those living animals.
Acknowledgements. I thank Kai Caspar for checking the text and providing useful suggestions and corrections.
For previous Tetrapod Zoology articles on dinosaur brains, biology and connected issues, see…
Could We Domesticate (Non-Bird) Dinosaurs?, August 2018
Postcranial Palaeoneurology and the Lifestyles of Pterosaurs, August 2018
Alternative Timeline Dinosaurs, the View From 2019 (Part 3): the Dinosauroid and its Chums, December 2019
Humanoid Dinosaurs Revisited Again: Russell and Séguin’s Dinosauroid at (Nearly) 40 Years Old, August 2021
A brain for Baryonyx: using CT-scanning to examine British spinosaurid brains, February 2023
T. rex the Tool Maker -- Testing Controversial Claims About Dinosaur Smarts, April 2024
Refs - -
Benton, M. J. 2021. The origin of endothermy in synapsids and archosaurs and arms races in the Triassic. Gondwana Research 100, 261-289.
Herculano-Houzel, S. 2023. Theropod dinosaurs had primate-like numbers of telencephalic neurons. Journal of Comparative Neurology 531, 962-974.
Osvath, M., Němec, P., Brusatte, S. L., & Witmer, L. M. 2024. Thought for food: the endothermic brain hypothesis. Trends in Cognitive Sciences 28, 998-1010.



