Well, here we are the final part in this long-running series. Thanks for sticking with it, and thanks for all the brilliant insight and annotation so many of you have provided in the comments (the series has been a real winner in terms of attracting visitors, and commenters especially).
Stop Saying That There Are Too Many Sauropod Dinosaurs, Part 7
Welcome to part – oh my god – seven in this seemingly eternal series.
Like me, I’m sure you want it to end so I can get back to writing about the innumerable other things on the list. Yes, we’re here, once again, for another instalment in the Too Many Damn Dinosaurs (TMDD) series. If you’re new to the whole thing, go back to Part 1 and see what this is all about; if you want to see all previous parts in the series go to the bottom of the article for the links (or use the sidebar). In the most recent articles, we looked at two assumptions inherent to the TMDD contention: that sauropod populations were similar in structure to modern megamammal populations, and that sauropods and other big dinosaurs were similar to Holocene megamammals in ecology and distribution. Here, we look at a third assumption, and it’s one that just won’t die.
Stop Saying That There Are Too Many Sauropod Dinosaurs, Part 6
Oh wow, we’re at Part 6 in the Too Many Damn Dinosaurs (TMDD) series already. You’ll need to have seen at least some of the previous articles to make sense of this one: you can either follow the links below, or click through the links in the sidebar. In Part 5 we looked at the first of a series of assumptions made by those who’ve advocated the TMMD contention; namely, that Late Jurassic sauropods had a population structure similar to that of megamammals. In this article, we look at a second assumption…
Stop Saying That There Are Too Many Sauropod Dinosaurs, Part 5
If you’ve been visiting TetZoo over recent weeks, you’ll know why we’re here. Yes, we’re here to continue with the Too Many Damn Dinosaurs (TMDD) series, in which I argue that it’s wrong to argue – that is, on principle, rather on detailed evaluation of the evidence – that the world famous Late Jurassic Morrison Formation contains too many sauropods. In the previous four parts of this series we introduced the DMDD contention, we looked at the fact that Paleogene mammals are not especially relevant to the TMDD contention, and then at the fact that modern giraffes are not especially relevant to the TMDD contention either.
Stop Saying That There Are Too Many Sauropod Dinosaurs, Part 4
In the previous articles in this series (see part 1 here, part 2 here and part 3 here) we looked at the ‘too many damn dinosaurs’ (TMDD) contention, this being the claim that the Upper Jurassic Morrison Formation simply has too many sauropod dinosaurs. You’ll need to check those previous articles out before reading this one. The previous parts of the series introduce the TMDD contention and then discuss whether arguments made about Paleogene fossil mammals and modern giraffes are relevant. Here, we move on to something else.
Stop Saying That There Are Too Many Sauropod Dinosaurs, Part 3
Welcome to the third part in this lengthy series of articles, all of which are devoted to the argument that those Mesozoic faunas inhabited by multiple sauropod taxa – in particular those of the Late Jurassic Morrison Formation – have too many damn dinosaurs (TMDD!). You need to have read parts 1 and 2 for this to make sense. Those articles set up the TMDD contention, and then showed why arguments relating sauropod diversity to Paleogene mammal diversity are erroneous. In this article, we look at another mammal-based argument.
Stop Saying That There Are Too Many Sauropod Dinosaurs, Part 2
A few authors would have it that there are too many damn dinosaurs (TMDD!): that the rich sauropod assemblage of the Late Jurassic Morrison Formation of the continental western interior of the USA simply contains too many species, and that we need to wield the synonymy hammer and whack them down to some lower number. In this article and those that follow it, I’m going to argue that this view is naïve and misguided. You’ll need to have read Part 1 – the introduction – to make sense of what follows here. Ok, to business…
Stop Saying That There Are Too Many Sauropod Dinosaurs, Part 1
Theropod Dinosaurs of the English Wealden, Some Questions (Part 1)
I have no idea whether I’m known for being a specialist on anything. But of the several zoological subject areas I publish on, among my favourite and most revisited is the dinosaurs of the English Wealden, and in particular the theropods (that is, the predatory dinosaurs) of the English Wealden.

Caption: at left, a Wessex Formation scene, depicting Eotyrannus, a compsognathid (at lower right), a pachycephalosaurian Yaverlandia in the middle distance, and the titanosauriform ‘Angloposeidon’. I need to do some new Wealden dinosaur artwork. At right: a younger, slimmer version of this blog’s author, holding the holotype claw of Baryonyx walkeri in 2001 or thereabouts. Images: Darren Naish.
What is the Wealden? It’s a Lower Cretaceous succession – formed of sandstones, siltstones, mudstones, limestones and clays – which was deposited during the Early Cretaceous, its oldest layers being from the Berriasian (and thus about 143 million years old) and its youngest from the early Aptian (and thus about 124 million years old). The sedimentology, subdivisions and terminology of the Wealden are complicated, but all you need to know here is that the whole lot is termed the Wealden Supergroup, that it has an old section called the Hastings Group and a younger section called the Weald Clay Group – both of which crop out on the English mainland – and that there’s also a young section called the Wealden Group that mostly crops out on the Isle of Wight. Finally, you also need to know that the Wealden Group includes the Wessex and Vectis formations. Yikes, even that brief summary was complicated, sorry.
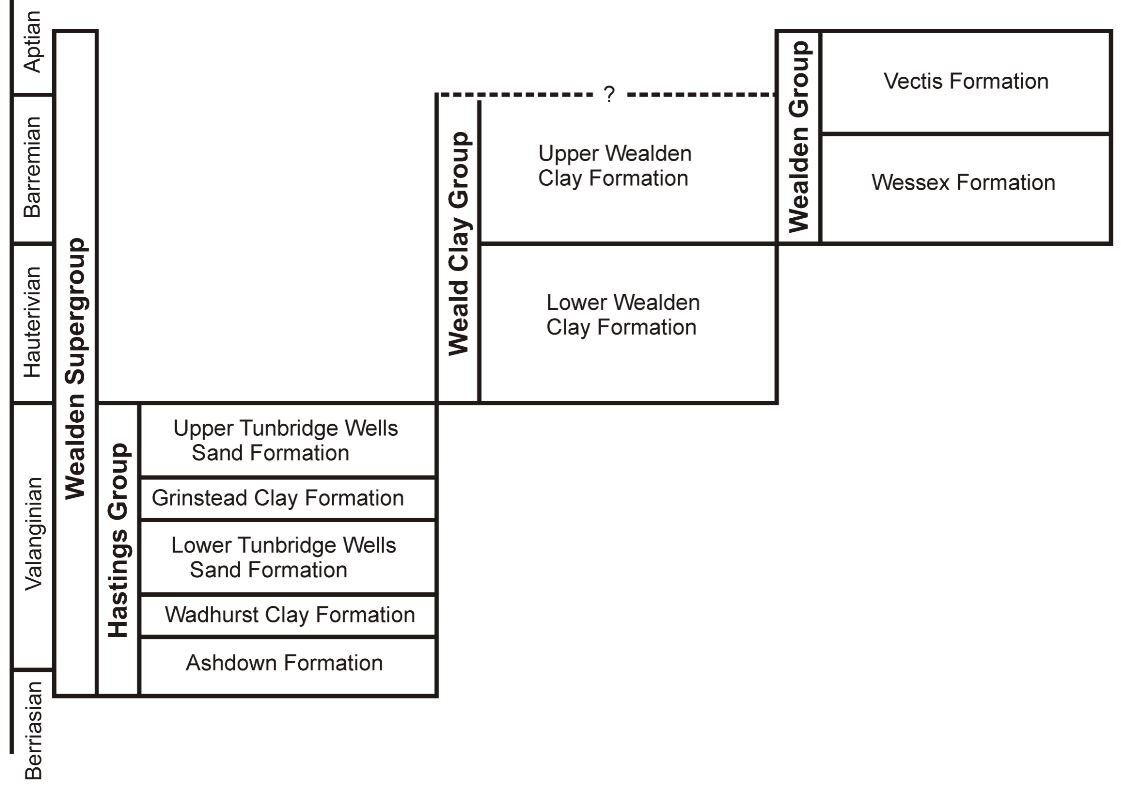
Caption: simplified stratigraphic nomenclature of the Wealden Supergroup. Note that the Hastings Group is much older than the Weald Clay and Wealden groups. The vast majority of Wealden dinosaurs come from the Wessex Formation. Image: Naish (2010).
The really interesting thing about the Wealden is that it’s highly fossiliferous, yielding everything from pollen and diatoms to dinosaurs. Wealden dinosaurs have been hugely important to our evolving understanding of these animals, in part because some of the earliest discoveries – Iguanodon, Hylaeosaurus and Hypsilophodon among them – come from the Wealden succession. Many Wealden dinosaurs have also been famously vexing, in part because they were discovered at a comparatively early stage in our knowledge, in part because their remains have been (and still are) highly incomplete, and in part because their historical taxonomy is a convoluted nightmare. Note also that the circa 20 million year duration of the Wealden means that its dinosaurs were not all contemporaries. Instead, they belonged to a series of distinct faunal assemblages. Within the last few decades, the Wealden has – focusing here on theropods alone – yielded the superstars Baryonyx, Neovenator and Eotyrannus, and its potential to give us really spectacular finds even today is affirmed by additional theropods that are yet to be published.

Caption: the Wessex Formation allosauroid Neovenator – here shown with some of its facial bones in partial x-ray – was covered here at TetZoo (ver 3) back in 2017. Our conclusions on the facial anatomy of this dinosaur (Barker et al. 2017) have since been challenged. Image: Darren Naish.
While I could say a whole lot more (I’ve co-authored a whole book on Wealden dinosaurs: Martill & Naish 2001), the point of the article here (and its follow-up, to be published later) is to provide a progress report of sorts on a few contentious or in-prep areas of Wealden theropod research. And I’ll admit right now that the topics I cover here are unashamedly based on my own research interests and projects, sorry. To work…

Caption: at left, Martill & Naish (2001) (cover art by Julian Hume). At right, Batten (2011), truly a must-have volume on Wealden palaeontology. Martill & Naish (2001) is now hard to get and only sold at ridiculous prices.
What are you, Yaverlandia? In 1923, Mr F. M.G. Abell discovered the partial skull roof of a fossil reptile at Yaverland on the Isle of Wight. Its thickened bone immediately led Watson (1930) to suggest that it might be from a pachycephalosaur. Fast forward now to the 1970s: Peter Galton – at the time, revising and redescribing all British ornithischians – took this idea and ran with it. He formally named the specimen Yaverlandia bitholus and argued that it was indeed a pachycephalosaur, the most archaic known (Galton 1971). This became the standard take on this dinosaur and the one I supported when writing Dinosaurs of the Isle of Wight in 2001 (Naish & Martill 2001).

Caption: at top, the Yaverlandia holotype in (left) ventral and (right) dorsal view. Below, the source of shame. Images: Darren Naish.
During my PhD years I was inspired to think about Yaverlandia anew, mostly because Jim Kirkland and Robert Sullivan (busy at the time with pachycephalosaurs) were pushing the idea that Galton’s identification was very likely wrong. I borrowed the specimen, produced a redescription, had the specimen CT-scanned, and photographed it to death. And I discovered a bunch of new stuff, all of which convinced me that Yaverlandia was not a pachycephalosaur at all, but a theropod. This data formed a chapter of my PhD thesis and brief summaries of my conclusions have been made here and there, including at conferences and in Naish (2011).

Caption: life reconstructions of Yaverlandia are few and far between. This one (seeming to show the animal in a quadrupedal pose: note how the artist has hidden the hand, a classic case of trying to cover up a mistake) is from the fabled Orbis part-work magazine series. I think (but can’t confirm) that the artist was Jim Channel. Image: (c) Orbis.
But the full, detailed explanation of the theropod hypothesis hasn’t yet appeared, though I promise that it will eventually (it’s a work I’m co-authoring with Andrea Cau). As is so often the case with my academic projects, I haven’t been able to make time to finish it (insert reminder about all my academic research being unfunded and done in ‘spare time’: I am not employed in academia). I should also add that a second specimen of Yaverlandia is known and also awaits writing-up. That’s a study I’m doing with Steve Sweetman.
![It’s well known in the theropod research community that the full description of this amazing fossil - the holotype of the Spanish ornithomimosaur Pelecanimimus - was done back in the 1990s [UPDATE: nope, 2004], but hasn’t seen print for a bunch of r…](https://images.squarespace-cdn.com/content/v1/510be2c1e4b0b9ef3923f158/1585002681161-29JO3RG3SRL02I363HHH/Wealden-theropod-questions-Mar-2020-Pelecanimimus-holotype-skull-1274px-250kb-Mar-2020-Darren-Naish-Tetrapod-Zoology.JPG)
Caption: it’s well known in the theropod research community that the full description of this amazing fossil – the holotype of the Spanish ornithomimosaur Pelecanimimus – was done back in the 1990s [UPDATE: nope, 2004], but hasn’t seen print for a bunch of reasons. Consequently, good information on the specimen isn’t (yet) available. Despite that, this photo has been widely shared online. I don’t know who to credit for it.
Are there ostrich dinosaurs in the Wealden or not? Back in the day, I was thrilled by the 1994 description of the remarkable multi-toothed ornithomimosaur Pelecanimimus polyodon from the Barremian Calizas de La Huérguina Formation of Spain. Not just because it’s a neat dinosaur, but because the Calizas de La Huérguina Formation has a lot in common with the Wealden: the two share a list of amphibians, mammals, lizards, crocodyliforms and dinosaurs, this rendering it plausible or likely that Pelecanimimus (or a similar taxon) might await discovery in the Wealden too (Naish et al. 2001, Naish 2002). Predicting the presence of a given group in a given faunal assemblage is a cheap and easy thing to do, and you can award yourself points for prescience and smarts if you’re proved right (even though most people will ignore your prediction), and no-one cares or notices if you never are. So, I’m not looking for a Wealden cookie here. Whatever, “where are the Wealden ornithomimosaurs?” was a question on my mind for a while.

Caption: these drawings – produced for Dino Press magazine back in 2002 – look very dated now. They’re supposed to show those smaller theropod groups confirmed for the Wealden (at top) and predicted for the Wealden but still awaiting discovery (at bottom). Images: Darren Naish.
So, I was pretty happy when – in 2014 – Ronan Allain and colleagues announced their discovery of such creatures in the Wealden. They’d discovered a new theropod in the Lower Cretaceous of France (specifically, in the Hauterivian or Barremian of Angeac in Charente, southwestern France) and had used this as a ‘Rosetta Stone’ in the interpretation of other Lower Cretaceous European theropod fossils (Allain et al. 2014). Several Wealden theropods – Valdoraptor, the Calamosaurus tibiae and Thecocoelurus among them – were ornithomimosaurs according to this study (Allain et al. 2014).

Caption: in 2014, I superimposed an ornithomimid into the Wessex Formation scene you saw above… this effort was not meant to be entirely serious (and an ornithomimid is the wrong kind of ornithomimosaur anyway). Image: Darren Naish.
I was initially enthusiastic about this proposal and thought that the authors were likely right. But as more and more information has been released on the Angeac theropod, the less like an ornithomimosaur it seems. It looks, instead, like a noasaur. Furthermore, the assorted relevant Wealden remains aren’t as similar to the bones of the Angeac animal as initially argued (Mickey Mortimer pointed this out in an article of 2014). Proper evaluation of what’s going on here will have to wait until a full description of the Angeac theropod appears in print. But if the Angeac theropod is a noasaur, the possibility that it’s close to or congeneric with one or more Wealden theropods remains a likelihood: Thecocoelurus (named for a single cervical vertebra from the Wessex Formation) looks like a noasaur vertebra (Naish 2011)... though that doesn’t necessarily mean that it is (since it also looks like an oviraptorosaur or therizinosaur vertebra in some features).

Caption: Mickey produced this image for a 2014 article at The Theropod Database (here).
To bring this round full circle, we might still be missing those predicted Wealden ornithomimosaurs.
Are there other Wealden tyrannosauroids besides Eotyrannus? Loooong-time readers of my stuff – I mean, those who’ve been visiting TetZoo since 2006 – might remember my suggestion from way back that some of the smaller theropod specimens from the Wealden are sufficiently similar to tyrannosauroids from the Lower Cretaceous of China to perhaps be additional small-bodied members of this group. I’m talking about Calamosaurus foxi (named for two cervical vertebrae), Aristosuchus pusillus (named for a partial pelvis and sacrum) and a few additional bits and pieces, including the so-called Calamosaurus tibiae (note the plural there). If these remains do belong to tyrannosauroids, they’re from taxa distinct from Eotyrannus (which everyone agrees is a tyrannosauroid).

Caption: the phylogeny I generated for my PhD thesis led me to think that Mirischia might be a tyrannosauroid… in which case Aristosuchus might also be a tyrannosauroid. This isn’t supported, however, in the in-prep Eotyrannus study I’ve co-authored with Andrea Cau. Image: Darren Naish.
I formally suggested a tyrannosauroid identity for Calamosaurus in a 2011 review of Wealden theropods (Naish 2011) but opted to keep Aristosuchus as a compsognathid on account of its similarity with Mirischia from Brazil. However, Mirischia also looks tyrannosauroid-like in some details (it has an anterodorsal concavity on the ilium) and I’ve sometimes wondered if it might also be a member of this clade. Recent results, however, do not support this possibility.

Caption: Aristosuchus pusillus is known from a sacrum and its conjoined pubic bones, which possess a notably long, narrow pubic boot (shown in ventral view in the image at bottom right). At left, we see where these bones would fit within the animal (here portrayed as a corpse; the reconstruction is dated and was produced for a conference poster I presented in 1999). Images: Darren Naish, Owen (1876).
So… Calamosaurus, are you a tyrannosauroid or not? When you only have two cervical vertebrae to go on (plus some tibiae that may or may not from the same taxon), it’s about impossible to say, and you can’t resolve things until you have better material. Like, an associated skeleton.

Caption: at left, one of the two holotype Calamosaurus foxi vertebrae in multiple views (from Naish et al. 2001). The bone is about 40 mm long in total. At right, a schematic reconstruction showing the two vertebrae in place in the cervical column of a compsognathid- or tyrannosauroid-like coelurosaur (from Naish 2002). Scale bar = 50 mm.
On that note, long-time readers might also recall my mention of a fairly good, associated skeleton of what appears to be a small Wealden tyrannosauroid. But it’s in private hands. I’ve been told by a British palaeontologist that the specimen concerned won’t be available for “this generation” of dinosaur specialists and I should simply forget about it. That’s hard, really hard.
And that’s where we’ll stop now. A second part to this article will be published soon.
For previous TetZoo articles on Wealden theropods and other dinosaurs, see (linking here to wayback machine versions due to destruction or paywalling of everything at versions 2 and 3)…
Of Becklespinax and Valdoraptor, October 2007
The world’s most amazing sauropod, November 2007
Oh no, not another new Wealden theropod!, June 2009
Concavenator: an incredible allosauroid with a weird sail (or hump)... and proto-feathers?, September 2010
The Wealden Bible: English Wealden Fossils, 2011, November 2011
Ostrich dinosaurs invade Europe! Or do they?, June 2014 (every archived version of this article lacks the original illustrations, sorry)
Refs - -
Allain, R., Vullo, R., Le Loeuff, J. & Tournepiche, J.-F. 2014. European ornithomimosaurs (Dinosauria, Theropoda): an undetected record. Geologica Acta 12, 127-135.
Galton, P. M. 1971. A primitive dome-headed dinosaur (Ornithischia: Pachycephalosauridae) from the Lower Cretaceous of England and the function of the dome of pachycephalosaurids. Journal of Paleontology 45, 40-47.
Naish. D. 2002. Thecocoelurians, calamosaurs and Europe’s largest sauropod: the latest on the Isle of Wight’s dinosaurs. Dino Press 7, 85-95.
Naish, D. 2010. Pneumaticity, the early years: Wealden Supergroup dinosaurs and the hypothesis of saurischian pneumaticity. In Moody, R. T. J., Buffetaut, E., Naish, D. & Martill, D. M. (eds) Dinosaurs and Other Extinct Saurians: A Historical Perspective. Geological Society, London, Special Publications 343, pp. 229-236.
Naish, D. 2011. Theropod dinosaurs. In Batten, D. J. (ed.) English Wealden Fossils. The Palaeontological Association (London), pp. 526-559.
Naish, D., Hutt, S. & Martill, D. M. 2001. Saurischian dinosaurs 2: Theropods. In Martill, D. M. & Naish, D. (eds) Dinosaurs of the Isle of Wight. The Palaeontological Association (London), pp. 242-309.
Naish, D. & Martill, D. M. 2001. Boneheads and horned dinosaurs. In Martill, D. M. & Naish, D. (eds) Dinosaurs of the Isle of Wight. The Palaeontological Association (London), pp. 133-146.
Owen, R. 1876. Monograph of the fossil Reptilia of the Wealden and Purbeck Formations. Supplement 7. Crocodilia (Poikilopleuron), Dinosauria (Chondrosteosaurus). Palaeontographical Society Monograph 30, 1-7.
Watson, S. 1930. Cf. Proodon [sic]. Proceedings of the Isle of Wight Natural History and Archaeology Society 1930, 60-61.
Minuscule Hummingbird-Sized Archaic Birds Existed During the Cretaceous
UPDATE (added 13th March 2020): since I published the article below, two relevant matters have come to attention, both of which have implications for the fossil discussed in the article.

Article at left from New Scientist; article at right from New York Times.
The first is that the extraction of amber from the locations concerned is linked with significant humanitarian issues. These make the continued publication and promotion of Burmese amber fossils look unethical; I was only dimly aware of these when writing the article and now regret my (minor) role in the promotion of this discovery (I did plan to delete the article but, on advice, was encouraged to keep it, but add the disclaimer you’re reading now). You can read about the humanitarian issues here, here and here.
Secondly, a number of experts whose opinions I respect have expressed doubts about the claimed theropod status of the fossil discussed below and have argued that it is more likely a non-dinosaurian reptile, perhaps a drepanosaur or lepidosaur (and maybe even a lizard).

A few artists have already produced speculative life reconstructions of Oculudentavis as a lepidosaur or similar reptile. It would have to be a big-brained, shallow-snouted, big-eyed one. Image: (c) Mette Aumala, used with permission.
I did, of course, consider this sort of thing while writing the article but dismissed my doubts because I assumed that - as a Nature paper - the specimen’s identity was thoroughly checked and re-checked by relevant experts before and during the review process, and that any such doubts had been allayed. At the time of writing, this proposed non-dinosaurian status looks likely and a team of Chinese authors, led by Wang Wei, have just released an article arguing for non-dinosaurian status. I don’t know what’s going to happen next, but let’s see. The original, unmodified article follows below the line…
—————————————————————————————————————————————
If you’ve been paying attention to 21st century palaeontological discoveries you’ll know that our understanding of Cretaceous vertebrate diversity has been much enhanced in recent years by the discovery of animals preserved within amber.

A number of really interesting vertebrate fossils in amber have been published in recent years. Among them are the two partial enantiornithine bird wings shown at left (both from Xing et al. 2016a), and the tiny anguimorph lizard Barlochersaurus winhtini (from Daza et al. 2018). Images: Xing et al. (2016), CC BY 4.0, Daza et al. (2018).
These include lizards and snakes (Daza et al. 2016, 2018), a segment of dinosaur tail originally identified as that of a non-bird theropod (Xing et al. 2016b), and various small birds (e.g., Xing et al. 2016a, 2017, 2018, 2019), all of which belong to the archaic, globally distributed group known as the enantiornithines or opposite birds. Today sees the latest of such discoveries, and it’s the most remarkable announced so far. It is – in fact – among the most remarkable of Mesozoic fossils ever announced, and I say this because of the implications it has for our understanding of Mesozoic vertebrate diversity.

Life reconstruction of Oculudentavis khaungraae Xing et al., 2020, depicting it as a tree-dwelling avialan theropod with partly colourful plumage. Image: (c) Gabriel Ugueto, used with permission.
The fossil in question, described in the pages of Nature by Lida Xing, Jingmai O’Connor and colleagues, is the complete, anatomically pristine but minuscule skull of a maniraptoran theropod – specifically, an archaic bird – named Oculudentavis khaungraae (Xing et al. 2020). The skull is preserved in a small amber block (31.5 x 19.5 x 8.5 mm) dating to the Cenomanian age of the Late Cretaceous (making it about 99 million years old). Like virtually all recently described amber vertebrates, it’s from Myanmar (Xing et al. 2020).

One of several images of the tiny Oculudentavis skull provided by Xing et al. (2020), this one (from their Extended Data) showing the specimen in left lateral view. The scale bar is 2 mm. Image: Xing et al. (2020).
When I say that this fossil is ‘minuscule’, I’m not kidding: the entire skull – the whole skull – is 14 mm long (1.4 cm; not a typo)*. This means that – at a very rough guess – the whole animal was around 90 mm (9 cm) long, an estimate I arrived it by producing a very schematic skeleton which equips the animal with a long tail. Xing et al. (2020) very rightly compare Oculudentavis with small hummingbirds: if it had a long bony tail (which it should have, given its inferred phylogenetic position; read on), it would have been longer than the tiny Mellisuga hummingbirds, the total lengths of which are around 50-60 mm, but not by much. It was unbelievably tiny.
* I’m frustrated by the fact that the authors don’t – so far as I can tell – provide the length of the entire skull anywhere in the paper, nor is there a table of measurements or an effort to estimate the animal’s complete size. Which is weird, because surely this is the most interesting thing about it.

A very rough, semi-schematic skeletal reconstruction of Oculudentavis which I produced in order to gain a rough idea of possible size. As you can see, it would have been tiny. The overall form of the skeleton is based on that of jeholornithiform birds; read on. Image: Darren Naish.
The skull of Oculudentavis has a typical ‘birdy’ look. It has a longish, shallow rostrum, large eye sockets, a lot of bone fusion (no, it isn’t a baby) and a rounded cranium where the section posterior to the eyes is short and compact (Xing et al. 2020). The nostrils are retracted, there’s no trace of an antorbital fenestra, the bony bars beneath the eye sockets bow outwards, and there’s a complete bony bar separating each eye socket from the openings at the back of the skull (Xing et al. 2020).
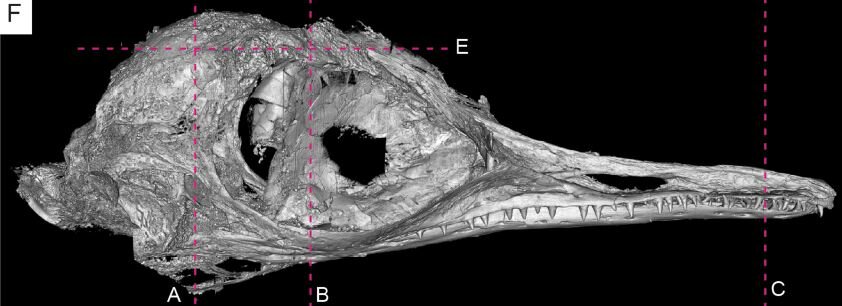
Digital scan of the skull of Oculudentavis in right lateral view (from the Extended Data of Xing et al. 2020). Note the overall toothiness. The dotted lines here show where slices were recorded during the scanning process. Image: Xing et al. (2020).
It’s a toothy little beast, with an atypically high number of conical (or near-conical) teeth lining its jaws all the way back to beneath the eye socket. This is unusual, since the toothrow in toothed birds and bird-like theropods in general normally stops well anterior to the eye. Another unusual feature is that the teeth aren’t located in sockets but are either fused to the jaw bones (the acrodont condition) or located within grooves that extend along the length of the jaws (the pleurodont condition) (Xing et al. 2020). The teeth look prominent, such that it’s hard to understand how they could be sheathed by lip tissue, nor is any such tissue preserved. Remember that beak tissue doesn’t occur in the same part of the jaws as teeth do, so Oculudentavis wouldn’t have had a true horny covering on its jaws. I assume that it had ‘lip’ tissue sheathing its teeth (except perhaps for the tips of the longest ones), as do other terrestrial tetrapods.

Speculative life reconstruction of Oculudentavis, its feathering and other details inspired by Jeholornis and other archaic members of Avialae. I’ve depicted it on the forest floor but am not necessarily saying that this is where it spent all of its time. Image: Darren Naish.
The eyes are directed laterally and the authors note that Oculudentavis likely didn’t have binocular vision (Xing et al. 2020). The sclerotic rings are huge and fill up most of the eye sockets. Xing et al. (2020) use the relative size of the eyes and their sclerotic rings to make inferences about the activity patterns and visual abilities of this animal: they think that Oculudentavis was likely day-active, had relatively small pupils, and perhaps had “unusual visual capabilities”.
The fossil doesn’t just consist of the animal’s bones alone, because synchrotron scanning reveals the presence of a brain (which is about as wide as it is long). Meanwhile, the bony palate preserves traces of its original tissue covering. This is decorated with numerous papillae, the first time such structures have been reported in a fossil theropod (Xing et al. 2020). The authors also refer to a tongue (!!) but it isn’t possible to make this out in the figures they provide, nor do they label it.
Combined, what do these features tell us about the lifestyle and ecology of Oculudentavis? The well-fused skull, prominent teeth and large eyes suggest that this was a predator, presumably of small arthropods. The soft papillae on the palate are of the wrong sort for fish-eating (Xing et al. 2020). Its tiny size and forest habitat imply that it was arboreal or scansorial – as suggested by Gabriel’s artwork above – but the animals that surround it in the cladogram are mostly terrestrial, so the possibility that it foraged in leaf litter or took regular trips to the forest floor are also conceivable, perhaps. Could it have been a predator of worms, molluscs or even tiny vertebrates, like a dinosaurian shrew?

Oculudentavis would have looked noticeably small relative to other Mesozoic birds, though not absurdly so. It’s compared here with Archaeopteryx (at upper left) and an assortment of others, most of which are enantiornithines. These illustrations are for my in-prep giant textbook project. Image: Darren Naish.
What sort of bird is Oculudentavis? For starters, it’s the presence of fused premaxillary and braincase bones, the position and size of the nostril, eye, postorbital region and domed cranium which strongly indicate that Oculudentavis is a member of Avialae, the bird lineage within Maniraptora (though note that the authors prefer the term Aves for said lineage). They included it within a phylogenetic analysis and found it to be one step more crown-ward (meaning, one node on the cladogram closer to living birds) than is Archaeopteryx, which is surprising because it makes Oculudentavis one of the most archaic members of the bird lineage (Xing et al. 2020). This could mean that birds underwent acute miniaturisation almost as soon as they evolved. Several authors – myself and colleagues included (Lee et al. 2016) – have argued beforehand that theropods on the line to birds underwent a gradual and pervasive decrease in size, but we didn’t (and couldn’t) predict that a size decrease of this sort occurred so early in bird history.

Theropods display a continuous, pervasive decrease in size when we look at the inferred size of ancestral species at successive nodes across the lineage leading to birds. From left to right, this illustration by Davide Bonnadonna shows the ancestral neotheropod (~220 Million years old), the ancestral tetanuran (~200 myo), the ancestral coelurosaur (~175 myo), the ancestral paravian (~165 myo), and Archaeopteryx (150 myo). Image: Davide Bonnadonna.
A World of Tiny Cretaceous Theropods? A key thing here is that we only know about this animal because of its preservation in amber: the rest of the fossil record mostly – the authors suggest – robs us of tiny vertebrates such as this. Could there actually have been many hummingbird-sized miniature theropods of this sort?

Xing et al. (2020) don’t provide a size estimate for Oculudentavis, but they do provide these silhouettes, which show Oculudentavis to scale with a hummingbird and chicken (and part of an ostrich is just visible at far right). Image: Xing et al. (2020).
Here’s where Xing et al.’s (2020) cladogram become especially interesting. The position they propose for Oculudentavis requires that its lineage originated about 150 million years ago, and yet Oculudentavis itself is about 99 million years old. Its lineage, therefore, is at least 50 million years long, in which case there could have been many of these tiny avialan dinosaurs (here, I have to resist the urge to talk about the hypothetical tree-climbing small dinosaurs of Dougal Dixon and George Olshevsky). I emphasise that this speculation assumes that the phylogenetic position Xing et al. (2020) infer is correct; it may not be. Indeed 10% of their trees found Oculudentavis in a different position: within enantiornithines, a possibility which seems ‘more right’ given the identity of other Burmese amber birds. With just a skull to go on, we obviously need more material before we can be especially confident on its phylogenetic position. And on that point, I won’t be surprised if it turns out that Oculudentavis does end up occupying a different position within maniraptoran theropods from the one which Xing et al. (2020) prefer. But none of this affects its minuscule nature, and that’s the real killer point here.

Part of a time-calibrated theropod tree (from Wang & Zhou 2017). According to Xing et al. (2020), Oculudentavis occupies a position more root-ward than Jeholornithiformes, but more crown-ward than Archaeopteryx. If correct, this means that its lineage originated during the latest part of the Jurassic. Image: Wang & Zhou (2017).
What About Other Fossil Vertebrates? If tiny, tiny Cretaceous theropods have remained unknown to us until now, what about other terrestrial vertebrates? I’d always assumed that the truly tiny frogs, lizards and other vertebrates of the modern world – those less than a few centimetres long – were recently evolved novelties of the Cenozoic. But maybe this is completely wrong. Maybe animals of this sort were present in the Mesozoic too, and maybe we’ve missed them due to a size filter which can only be filled by fossils discovered in amber?

The modern world is inhabited by truly tiny lizards and frogs, like this c 3cm SVL Brookesia chameleon and c 1cm Stumpffia frog. Were similarly tiny tetrapods also around in the Cretaceous? Images: (c) Mark D. Scherz, used with permission.
Time will tell. This is really exciting stuff.
For previous TetZoo articles relevant to this one, see…
Bird behaviour, the ‘deep time’ perspective, January 2014
50 million years of incredible shrinking theropod dinosaurs, July 2014
The Romanian Dinosaur Balaur Seems to Be a Flightless Bird, June 2015
The Most Amazing TetZoo-Themed Discoveries of 2018, December 2018
A Multi-Species Nesting Assemblage in the Late Cretaceous of Europe, February 2019
Refs - -
Daza, J. D., Bauer, A. M., Stanley, E. L., Bolet, A., Dickson, B. & Losos, J. B. 2018. An enigmatic miniaturized and attenuate whole lizard from the mid-Cretaceous amber of Myanmar. Breviora 563, 1-18.
Daza, J. D., Stanley, E. L., Wagner, P., Bauer, A. M. & Grimaldi, D. A. 2016. Mid-Cretaceous amber fossils illuminate the past diversity of tropical lizards. Science Advances 2 (3), e1501080.
Wang, M. & Zhou, Z. 2017. The evolution of birds with implications from new fossil evidences. In Maina, J. N. (ed) The Biology of the Avian Respiratory System. Springer International Publishing, pp. 1-26.
Xing, L., McKellar, R. C., O’Connor, J. K., Bai, M., Tseng, K. & Chiappe, L. M. 2019. A fully feathered enantiornithine foot and wing fragment preserved in mid-Cretaceous Burmese amber. Scientific Reports 9, 927.
Xing, L., McKellar, R. C., Wang, M., Bai, M., O’Connor, J. K., Benton, M. J., Zhang, J., Wang, Y., Tseng, K., Lockley, M. G., Li, G., Zhang, Z. & Xu, X. 2016a. Mummified precocial bird wings in mid-Cretaceous Burmese amber. Nature Communications 7, 12089.
Xing, L., McKellar, R. C., Xu, X., Li, G., Bai, M., Scott Persons IV, W., Miyashita, T., Benton, M. J., Zhang. J., Wolfe, A. P., Yi, Q., Tseng, K., Ran, H. & Currie, P. J. 2016b. A feathered dinosaur tail with primitive plumage trapped in mid-Cretaceous amber. Current Biology 26, 3352-3360.
Xing, L., O’Connor, J. K., McKellar, R. C., Chiappe, L. M., Bai, M., Tseng, K., Zhang, J., Yang, H., Fang, J. & Li, G. 2018. A flattened enantiornithine in mid-Cretaceous Burmese amber: morphology and preservation. Science Bulletin 63, 235-243.
Xing, L., O’Connor, J. K., McKellar, R. C., Chiappe, L. M., Tseng, K., Li, G. & Bai, M. 2017. A mid-Cretaceous enantiornithine (Aves) hatchling preserved in Burmese amber with unusual plumage. Gondwana Research 49, 264-277.
Xing, L., O’Connor, J. K., Schmitz, L., Chiappe, L. M. McKellar, R. C., Yi, Q. & Li, G. 2020. Hummingbird-sized dinosaur from the Cretaceous period of Myanmar. Nature 579, 245-249.
Did Dinosaurs and Pterosaurs 'Glow'? Extinct Archosaurs and the Capacity for Photoluminescent Visual Displays
One of many exciting discoveries made in tetrapod biology in recent decades is that UV-sensitive vision is not just a thing that exists, but a thing that’s widespread.

Caption: would a live dinosaur - like this heterodontosaur - look utterly different if its tissues were photoluminescent? Brian Engh explored this possibility in this excellent piece of art, included in Woodruff et al. (2020). Image: Brian Engh.
We’ve known since the early 1980s that at least some birds can detect UV wavelengths, and research published more recently has demonstrated its presence in lizards of disparate lineages, in turtles, rodents and, most recently, amphibians. Some of these animals use their UV-sensitive vision to find food (like pollen-rich flowers) and perhaps even to navigate their environments (UV-sensitive vision in certain forest-dwelling birds might enhance their ability to see certain kinds of leaves, for example).
That’s great, but what’s even more surprising – though maybe it shouldn’t be – is that markings and tissue types in some of these animals are visible to other animals with UV-sensitive vision. Furthermore, some tissue types are able to absorb UV and re-emit it within part of the spectrum visible to we humans. It’s this aspect of the UV story – the possibility that UV is absorbed and emitted as visible light (typically blue light) – that we’re talking about hereon, not UV-sensitive vision. Note that the terms used for this phenomenon are slightly contentious among relevant experts. Most agree that the right term is fluorescence whereas others (including my colleague Jamie Dunning) argue that we should use the more specific photoluminescence. I have no proverbial dog in this fight but am going to stick with photoluminescence here seeing as it’s the one we used in the relevant paper.

Caption: in 2018, Jamie Dunning and colleagues reported the discovery of photoluminescence in puffins. Image: (c) Jamie Dunning.
The discovery of photoluminescence in animals is evidently of broad general interest, and I can make this assertion because several recent studies reporting its occurrence have received an unusual amount of public interest. Dunning et al.’s (2018) report on its occurrence in the brightly coloured bill plates of puffins, for example, proved a really popular discovery (Wikinson et al. (2019) followed up with a subsequent study on the keratinous horns of rhinoceros auklets), as did Prötzel et al.’s (2018) discovery of photoluminescent bones in chameleons. Remarkably, Prötzel et al. (2018) were able to show that the ‘glowing’ bones of these lizards are visible through the skin. At the time of writing, a study reporting widespread photoluminescence in living amphibians has just appeared, and it too has received a fair amount of general interest.
Here it’s worth making a critical point on the popularity of these studies in the popular media. There’s no doubt that this stuff is interesting, and certainly of relevance to biologists at large (for one thing, knowing about the distribution of fluorescence/photoluminescence could have all kinds of implications for surveying and collecting). But there’s concern that the studies are being framed in the wrong way, and that more thorough vetting is needed, in places. Also worth noting is that what role photoluminescence actually has to the animals that emit it is controversial, since some workers argue (a) that its visual signalling role hasn’t been sufficiently tested for, and (b) it may simply be too subtle to be of much use to the animals in which it’s present. Keep this in mind when reading the following!

Caption: Prötzel et al.’s (2018) bone-glow research on chameleons shows that the photoluminescing bones of these lizards were actually visible through the skin. Image: David Prötzel.
These caveats notwithstanding, if UV-themed visual displays are widespread in tetrapods, those of us interested in fossil animals are presented with an interesting set of possibilities. We already think that the many extravagant structures of non-bird dinosaurs and pterosaurs – they include cranial horns, crests and casques as well as spikes, spines, sails, bony plates and so on – functioned predominantly in visual display. Could they also have been photoluminescent, and could this have then been used to enhance the display function of the structures in question?
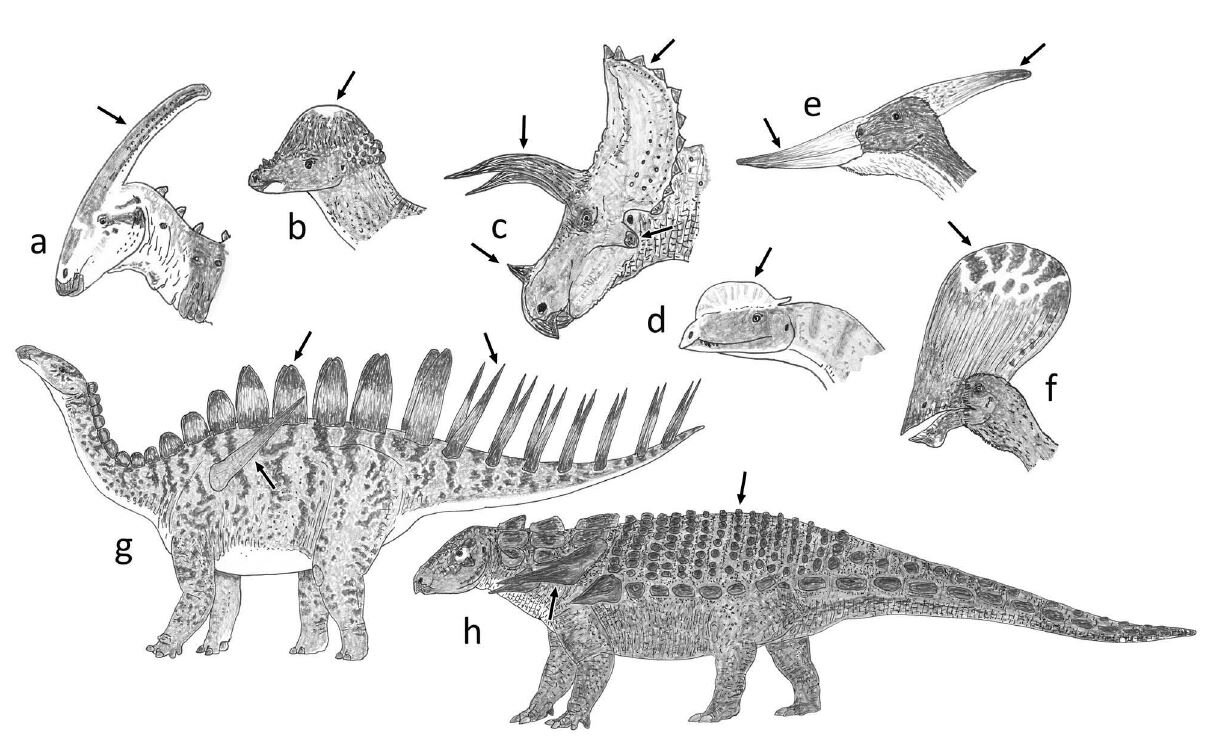
Caption: dinosaurs and pterosaurs are of course notable for their remarkable variety of what I term extravagant structures, a selection of which are depicted here. (a) Parasaurolophus, a hadrosaurid ornithopod. (b) Pachycephalosaurus. (c) Triceratops, a ceratopsid ceratopsian. (d) Dilophosaurus, a theropod. (e) Pteranodon and (f) Tupandactylus the pterodactyloid pterosaurs. (g) Miragaia the stegosaur. (h) Edmontonia the nodosaurid ankylosaur. From Woodruff et al. (2020), images by Darren Naish.
In a brand-new paper published this week in Historical Biology (or on its website, anyway), Cary Woodruff, Jamie Dunning and I set out to consider this very question (Woodruff et al. 2020). At the risk of spoiling the surprise I’ll say that we don’t provide a hard or definitive answer; our aim instead is to bring attention to the possibility that photoluminescence might have been present in some of these animals. We encourage the testing of this possibility and suggest some specific ways in which this testing might be performed. Of incidental interest is that our collaboration evolved from a Twitter discussion (which is currently findable here).

Caption: a palaeontologist ponders new papers on photoluminescence, and then gets talking to one of the relevant researchers. And I chimed in as well, sorry. The rest is history…
I should also add that our idea isn’t especially new. Ever since UV-sensitive vision was first reported in birds back in the 1980s, the idea that extinct dinosaurs might have made use of photoluminescence has been mooted (though, let me make the point again: you don’t need UV-sensitive vision to see photoluminescence). I’ve incorporated photoluminescence into more than one dinosaur-themed media project, most recently Dinosaurs in the Wild.
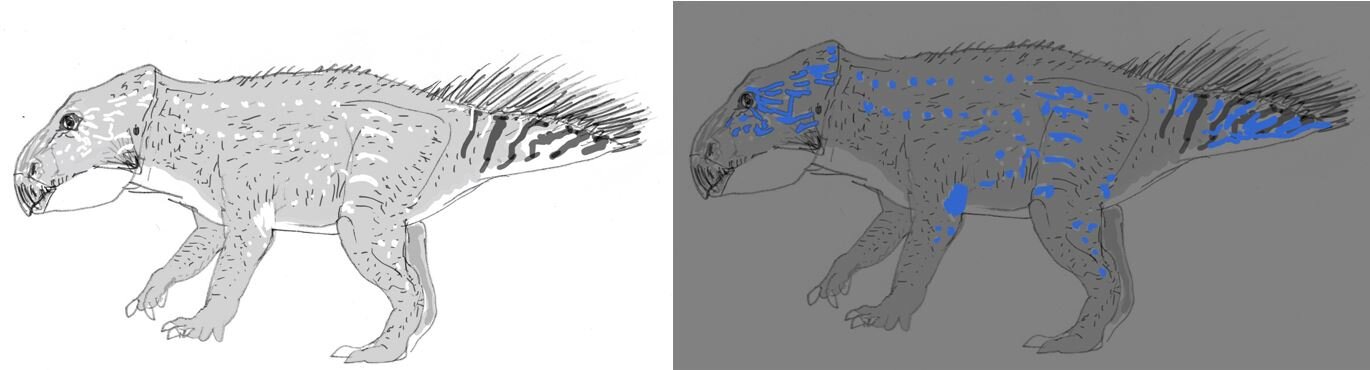
Caption: the idea that Mesozoic dinosaurs might have been exploiting photoluminescence isn’t altogether new. Here are rough sketches I produced depicting the concept of a photoluminescent Leptoceratops produced for the travelling visitor experience Dinosaurs in the Wild. Image: Darren Naish.
A few specific points are worthy of attention. Above, I mentioned Prötzel et al.’s (2018) chameleon-themed ‘bone glow’ study. Bone-based photoluminescence has also been reported in frogs, specifically in the Brachycephalus pumpkin toadlets (Gouette et al. 2019). Could those dinosaurs superficially similar to chameleons (namely ceratopsians: like some chameleons, they have bony frills and prominent horns) also possess bone-based photoluminescence and, if so, could they exploit it in chameleon-like fashion? Well, probably not, mostly because the much larger size of these dinosaurs means that their skin was too thick for this to work (Woodruff et al. 2020).

Caption: for fun, let’s use toy ceratopsians rather than the real things. Could these dinosaurs have had ‘glowing’ bones as modern chameleons do? No, almost certainly not. Image: Darren Naish.
One of the most unusual things about non-bird dinosaurs possessing extravagant structures is that males and females are extremely similar (albeit not necessarily identical) with respect to the form and proportional size of said structures. As regular TetZoo readers might recall from several articles published here within recent years (see links below), some workers interpret the extravagant structures of Mesozoic dinosaurs as functioning within a model of species recognition. According to this model, the structures function as banners used to signal membership of whatever the respective species is. I don’t think that this is valid for a bunch of reasons and in fact I don’t think that extravagant structures have an important role in species recognition at all (Hone & Naish 2013, Knell et al. 2013). An alternative model posits that extravagant structures mostly have an intraspecific function, work as sociosexual signals of reproductive quality, and evolved within the context of sexual selection. This is the model that I and my colleagues support (Hone et al. 2011, Knell et al. 2012, 2013, Hone & Naish 2013), and a lengthy debate that’s been thrashed out in the literature over the past decade pits species recognition and sexual selection as opposing schools of thought.

Caption: at left, mutual sexual selection at play in the Great crested grebe, as illustrated by Julian Huxley in 1914. At right, cover of the famous issue of TREE which includes Knell et al.’s (2012) seminal review.
But if this is so, why is it that ostensible males and females in the dinosaur species concerned are monomorphic: that is, they have similar extravagant structures? Back in 2011, Dave Hone, Innes Cuthill and I argued that these animals might have evolved their extravagant structures within the context of mutual sexual selection (Hone et al. 2011), this being the strategy where both males and females use their extravagant structures in sociosexual display. But while we know that extant monomorphic animals really are monomorphic, we’re not sure that this is (or was) the case for extinct ones: it could still be that their structures differed in hue, colour or some other visual property. If we’re speculating about the possible presence of photoluminescence in extinct archosaurs, the possibility exists that “monomorphic elaborate structures in pterosaurs and non-bird dinosaurs were not monomorphic in life” but differed in how they photoluminesced (Woodruff et al. 2020, p. 5). We were inspired by the sexually dimorphic photoluminescence of chameleons and Brachycephalus frogs.

Caption: could the in-situ, fully intact armour of ankylosaurs like that of the amazing holotype of Borealopelta, shown here, give insight into the potential of photoluminescence in these animals? Image: CC SA 4.0, original here.
Finally… speculating about the presence of photoluminescence is all very well and good, but can we test for it? In those cases where part of the integument is preserved, we can, by shining blacklights at the respective specimens. The problem, however, is that we might not be seeing the original light-emitting properties of the animal. Seemingly positive results might be a consequence of the fact that various tissues (bone included), minerals and preservatives fluoresce under UV (Woodruff et al. 2020).
As a preliminary test, we looked at the osteoderms of the spectacularly preserved ankylosaurs Borealopelta and Zuul under UV light… we did get results, but it’s difficult to know what, if anything, these results tell us about any condition present in life (Woodruff et al. 2020). I should add that people have been shining blacklights at fossils for a long time and seeing all kinds of interesting results (hat-tip to the pioneering work of Helmut Tischlinger); in no way are we implying that we’re anything like the first to do this.

Caption: people have been examining fossils with UV light for decades. These images show the Jurassic pterosaur Bellubrunnus roethgaengeri, illuminated via the use of UV. Image: Hone et al. 2012 (original here).
And that about wraps things up for now. As will be clear, our paper is not much more than a preliminary set of speculations and suggestions for further work, and isn’t intended to be an in-depth analysis of the proposal. But – as I see it – that’s ok: the scientific literature really shouldn’t be considered focused on results alone, since review, discussion and valid speculation are valuable and worthy too. I hope you agree.
UPDATE (adding 4th March 2020): this article has been somewhat modified relative to its original version, since a misunderstanding on my part meant that I was previously describing photoluminescence as a phenomenon especially relevant to animals with UV-sensitive vision. Substantial thanks to Michael Bok for his interest and assistance and for sending comments which enabled me to modify the article.
For previous TetZoo articles on the biology and life appearance of Mesozoic dinosaurs and pterosaurs, see (as usual now, linking to wayback machine versions due to vandalism and paywalling of ver 2 and 3)…
Zuniceratops and the early acquisition and alleged dimorphism of ceratopsian brow horns, April 2009
Necks for sex? No thank you, we’re sauropod dinosaurs, May 2011
Did dinosaurs and pterosaurs practise mutual sexual selection?, January 2012
Sexual selection in the fossil record, September 2012
Dinosaurs and their ‘exaggerated structures’: species recognition aids, or sexual display devices?, April 2013
Refs - -
Dunning, J., Diamond, A. W., Christmas, S. E., Cole, E. L., Holberton, R. L., Jackson, H. J., Kelly, K. G., Brown, D., Rojas Rivera, I. & Hanley, D. 2018. Photoluminescence in the bill of the Atlantic Puffin Fratercula arctica. Bird Study 65 (4), 1-4.
Goutte, S., Mason, M.J., Antoniazzi, M.M., Jared, C., Merle, D., Cazes, L., Toledo, L.F., el-Hafci, H., Pallu, S., Portier, H., Schramm, S., Gueriau, P. & Thoury, M. 2019. Intense bone fluorescence reveals hidden patterns in pumpkin toadlets. Scientific Reports 9, 5388.
Prötzel, D., Heß, M., Scherz, M. D., Schwager, M., van’t Padje, A. & Glaw, F. 2018. Widespread bone-based fluorescence in chameleons. Scientific Reports 8, 698.
Wilkinson BP, Johns ME, Warzybok P. 2019. Fluorescent ornamentation in the Rhinoceros auklet Cerorhinca monocerata. Ibis 161, 694-698.
Beautiful, Big, Bold Dinosaur Books: of Molina-Pérez and Larramendi’s Theropods, Rey’s Extreme Dinosaurs 2, and Parker et al.’s Saurian
One of the reasons you read TetZoo is because of the dinosaurs, and among the dinosaur-themed things I write about on fairly regular basis are new(ish) dinosaur-themed books.

Partly because I’m way overdue on the book reviews I planned to write during 2019, I’m here going to talk about some recently-ish published dinosaur-themed books that you’d do well to buy and read, if you wish, or can. I’ve written about recently-ish published dinosaur-themed books on quite a few recent occasions; see the links below for more. Let’s get to it.
Encyclopedia of Dinosaurs: the Theropods, by Rubén Molina-Pérez and Asier Larramendi

I can say right out of the gate that this 2019 work is one of the most spectacular dinosaur-themed works that has ever seen print. Think about that for a minute, since it’s a pretty grand claim. Yes. This book is spectacular: big (288 pages, and 24.5 cm x 30 cm), of extremely high standard, packed with information, and containing a vast number of excellent and highly accurate colour life reconstructions. Originally published in Spanish, it has now been translated (by David Connolly and Gonzalo Ángel Ramírez Cruz) and published in English by London’s Natural History Museum. The book consists of eight sections, which variously go through the theropod cladogram, discuss geographical regions and the theropods associated with them, and review theropod anatomy, eggs, footprints and so on. And it’s packed with excellent illustrations… hundreds of them.

A selection of pages from Molina-Pérez & Larramendi (2019). At left, eggs depicted to scale (with a basketball). At right, just two of the many pages that feature theropod skeletal elements. Images: Molina-Pérez & Larramendi (2019).
The art is great – the majority of colour images being by the phenomenally good and reliable Andrey Atuchin – and I’d recommend that anyone interested in the life appearance of dinosaurs obtain the book for its art alone. I have one criticism of the art though, which is that the colour schemes and patterns used for some of the animals are occasionally based on those of living animals (most typically birds).

Just two of the many UNNAMED theropod species reconstructed in the book. Exciting stuff! The humans that feature in the book are an interesting lot. Images: Molina-Pérez & Larramendi (2019).
Familiar theropods of many sorts are illustrated, but a major plus point is that many of the animals depicted are either obscure and mostly new to the world of palaeoartistic depictions (examples: Spinostropheus, Dryptosauroides, Bonapartenykus, Nanantius, Gargantuavis) or are as-yet-unnamed species: animals which clearly represent something new (since they’re the only member of their group known from the relevant geographical region and segment of geological time) but have only been referred to by their specimen numbers or by a ‘cf’ attribution (a theropod called, for example ‘cf Velociraptor mongoliensis’ is being compared by its describers to V. mongoliensis and is clearly very much like V. mongoliensis, but quite possibly not part of the species and perhaps something new. The ‘cf’ is short for ‘confere’, as in: compare with).

If the reconstructions in the book are anything to go by, Spinostropheus - according to one specimen (an ulna) it could reach huge sizes - was among the most remarkable of theropods. Just look at it. Image: Molina-Pérez & Larramendi (2019).
Two issues strike me as problematic though. One is that the arrangement is really difficult to get to grips with, and it’s taken me numerous attempts to understand and appreciate why the book is arranged the way it is. The second issue concerns what appears to be a suspiciously high degree of taxonomic precision for footprints. The authors depict footprints, said to be representative of the different theropod groups covered in the taxonomy section at the start of the book, and seem confident that the tracks concerned (which are often fairly nondescript) were made by species belonging to the relevant group. It’s hard to be convinced that this is reliable, except in a very few cases: I’m happy to agree that the giant Tyrannosauripus pillmorei track, for example, really was made by a member of Tyrannosauridae. Then again, maybe the authors have devised a new track identification method that isn’t yet known to the rest of us.
The several montages in the book are truly things of beauty. Image: Molina-Pérez & Larramendi (2019).
And on that note, it’s obvious that a vast quantity of novel science was performed as part of the background research for this book. The methods and data used by the authors are explained up front. Impressive stuff, and stuff which should be published in the technical literature at some point. Are their results always ‘good’? Well, I have my misgivings about the idea that Dinornis the moa was the fastest (non-flying) theropod ever and capable of sprinting at a phenomenal 81 km/h…

So, so many diagrams of tracks and trackways. Image: Molina-Pérez & Larramendi (2019).
Despite my minor misgivings, this book is attractive enough and interesting enough that it’s a must-have for those seriously interested in dinosaurs and in artistic depictions of them. Buy it if you can. This volume promises to be the first in a series. At the time of writing, the second of these – devoted to sauropods – is being advertised and is due to appear soon. Given that there’s every reason to assume that its artwork and overall quality will be similar to that of the theropod volume reviewed here, I have very high hopes and look forward to seeing it.
Molina-Pérez, R. & Larramendi, A. 2019. Encyclopedia of Dinosaurs: the Theropods. The Natural History Museum, London. pp. 288. ISBN 9780565094973. Hardback. Here at publishers. Here at amazon. Here at amazon.co.uk.
Luis Rey’s Extreme Dinosaurs 2: the Projects

Luis Rey (who blogs here) has been active in the palaeoart world for a few decades now, and most people interested in the portrayal of dinosaurs in art will be familiar with his vibrant, bold and dynamic style. Luis’s work has appeared in museum installations, exhibitions, and in numerous publications, including books. Most notable among these are his own Extreme Dinosaurs (Rey 2001) and The Big Golden Book of Dinosaurs, by Robert Bakker (Bakker 2013). I feel it would be wrong at this point to avoid mentioning the fact that Luis and I were regular correspondents back when the first of those books appeared, but that my dislike of the 2013 book – which I made clear in a TetZoo review – coincided with a cessation in any contact we used to have. But things have moved on; Luis’s art has continued to evolve and let’s put all of that behind us.

A collection of Rey works at TetZoo Towers. Image: Darren Naish.
Extreme Dinosaurs 2: the Projects discusses the intellectual and artistic background to several dinosaur-themed museum installations which Luis has created, but does so in an evolutionary fashion such that they’re used to describe our improving knowledge of the Mesozoic world. Luis was illustrating colourful, fully feathered dromaeosaurs, oviraptorosaurs and so on at a time when the majority of relevant academics were dead against this, so it would be fair to see him as one of several artists who were predicting things that would prove correct in the end. I was on side too, and consequently was a vociferous Rey advocate in the early part of my academic career, deliberately using his reconstructions of dromaeosaurs – even the turkey-wattled, shaggily feathered ones – in conference presentations and publications. And for all the success of All Yesterdays and its associated movement, we have never forgotten that Luis was saying many of the same things already.

Some representative pages from Extreme Dinosaurs 2. At right, note the person wearing an oviraptorosaur costume while sat in a nest.
Maniraptorans, thyreophorans, ceratopsians, dinosaur eggs and nesting and the dinosaurs (and other Mesozoic reptiles) of Mexico all get coverage here as Luis explains the thinking behind new pieces of art and also how and why he’s modified older ones. Our understanding of feather arrangements in non-bird maniraptorans have improved a lot in recent years, plus we have so much new data on ankylosaur armour, ceratopsian skin and so on. If you haven’t been keeping up, this book would be a good primer. The text is concise and written in a friendly, informal style.

More representative pages, this time depicting therizinosaurs. Brightly coloured faces, bold patterns on the feathering, inflatable throat structures… what’s not to love? Image: Rey (2019).
I’m still not keen on the photo-bashing that’s now integral to the Rey style and don’t find it effective or successful. A few pieces included in the book don’t, therefore, work for me (examples: Santonian hadrosaur hassled by theropods on pp. 104-5, the Labocania scene on pp. 106-7). But can I please emphasise that I still find this a valuable book, and I very much recommend it as an interesting addition to the palaeoart stable, the autobiographical angle in particular being useful. My copy of Extreme Dinosaurs 2: the Projects is an advance softback version but I understand that a hardback is available too.
Rey, L. 2019. Extreme Dinosaurs 2: the Projects. Imagine Publishing, London/Metepec. pp. 139. ISBN 978-0-9933866-2-6. Softback/Hardback. Here at amazon.co.uk.
Parker et al.’s Saurian: A Field Guide to Hell Creek

Even if – like me – you’re not a video game buff and have little interest in video games or the playing of them, chances are high that you’ve heard about Saurian, a role-playing, survival simulation experience in which you play the role of a dinosaur negotiating a Maastrichtian environment modelled on that of the Hell Creek Formation. You have to avoid predators, find and procure food, raise babies, and live as long as possible. I’ve played The Simms, back in the day, and recall the urge to sink hours of time into living a virtual tiny life, so I can understand the appeal.

Representative pages, here showing the raptor prey restraint model in action. Poor pachycephalosaur. Image: Parker et al. (2019).
The relevance of Saurian to the TetZooniverse is that the game has been designed and built according to an incredibly high scientific standard, the team behind it having done a vast quantity of research on everything relevant to the Hell Creek world. And the good news is that this work hasn’t been wasted. This book – written by Tom Parker, featuring the artwork of Chris Masna and RJ Palmer (of Detective Pikachu fame and much else), and produced following consultation with a long list of relevant palaeontologists – is the result: it includes a vast quantity of amazing concept art, detailed vignettes and scaled artwork of the organisms and environments that feature in the game, and is simply a joy to look at. The text is brief but functions well. The book is a sturdy softback (20 x 24 cm) of 182 pages, printed to excellent, glossy standard.

RJ Palmer’s T. rex is one of the stars of the show. Some of you will know that the Saurian team abandoned an earlier, more feathery version. Assorted humans function as scale bars; you can likely guess who this one is based on. Image: Parker et al. (2019).
As should be clear by now, the thoroughness of the project’s world-building means that there’s more information here on Maastrichtian North American trees, rainfall patterns, swamps, beaches, amphibians, lizards, crocodyliforms, fishes and so on than you’ve ever seen before. The result is one of the most interesting, detailed and attractive volumes dedicated to Late Cretaceous life. This book is a must-have for those seriously interested in palaeoart and in seeing prehistoric animals and environments depicted in detail, but it’s also good enough that those with a scientific or technical interest in Late Cretaceous life should obtain it too.

Oh wow, so much of the art in this book is just phenomenal. This scene depicts competition among scavengers at a carcass. You might just be able to see the anguimorph lizard inside the body cavity. Image: Parker et al. (2019).
Parker, T., Masna, C. & Palmer, R. J. 2019. Saurian: A Field Guide to Hell Creek. Urvogel Games. pp. 182. Softback. Here at publishers [BUT CURRENTLY OUT OF STOCK].
That’s where we’ll end for now. A few more dinosaur-themed book review are due to appear here soon, including of Donald Prothero’s The History of Dinosaurs in 25 Fossils and Michael Benton’s The Dinosaurs Discovered: How a Scientific Revolution is Rewriting Their Story.
For previous TetZoo dinosaur-themed book reviews, see… (linking here to wayback machine versions due to paywalling and vandalism of TetZoo ver 2 and 3 articles)…
Long and Schouten's Feathered Dinosaurs, a review, January 2008
Luis Chiappe's Glorified Dinosaurs: The Origin and Early Evolution of Birds, January 2011
What they’re saying about The Great Dinosaur Discoveries, November 2011
Greg Paul’s Dinosaurs: A Field Guide, August 2012
New Dinosaur Books, Part 1: Barrett on Stegosaurs, August 2018
New Dinosaur Books, Part 2: Ben Garrod’s ‘So You Think You Know About… Dinosaurs’ Series, August 2018
The Second Edition of Naish and Barrett’s Dinosaurs: How They Lived and Evolved, November 2011
Refs - -
Bakker, R. T. 2013. The Big Golden Book of Dinosaurs. Random House, New York.
Rey, L. 2001. Extreme Dinosaurs. Chronicle Books, San Francisco.
Philip J. Senter’s Fire-Breathing Dinosaurs?, the TetZoo Review
Many of us interested in the more arcane side of natural history will be aware of that body of literature that seeks to explain the biology, behaviour and history of living things within the words of a complex, multi-authored work known as The Bible. I refer of course to the creationist literature; to that number of books and articles whose authors contend that animals known from fossils simply must accord with the stories and descriptions of the Bible, and whose authors furthermore contend that the Earth and its inhabitants must have come into being within the last few thousand years.

Caption: a smouldering Parasaurolophus: the cover art for the book, by Leandra Walters. Image: (c) Leandra Walters/Senter (2019).
Creationist authors – the most familiar include Ken Ham, Kent Hovind and Duane Gish – have argued that non-bird dinosaurs and other fossil animals were inhabitants of the Garden of Eden, that predatory species like Tyrannosaurus rex ate water melons and sugarcane before The Fall, that humans and animals like Tyrannosaurus lived alongside one another during the early days of the Earth’s creation, that evolution cannot have happened, except when it did as species emerged from their different ancestral kind (or baramins), and that animals like Tanystropheus, tyrannosaurs and pterosaurs were seen and written about by people, and are responsible for the mythological creatures mentioned or described in the Bible and other ancient texts. Leviathan and Behemoth of The Bible, Grendel of the medieval epic Beowolf, the fire-breathing dragons of the Middle Ages and so on must – creationist authors contend – be descriptions of human encounters with giant reptiles otherwise known as fossils. And, yes: you read that right… creationist authors have argued, apparently seriously, that fire-breathing dragons must be descriptions of encounters with animals like dinosaurs and pterosaurs. So… they… breathed fire, then.

Caption: the Bible specifically states that the first few books of the Old Testament are not meant to be taken literally. Despite this, a number of Young Earth creationists promote a view of the ancient world where people lived alongside allosaurs and pterosaurs and so on. If you’ve seen a version of this page mentioning lemonade and homosexuality, it’s a spoof (the original text does not include that section of text). Image: (c) Ken Ham, Dinosaurs of Eden.
Over the past several years, Fayetteville State University biologist and palaeontologist Philip J. Senter has published a great many technical scientific articles evaluating the various claims and models of creationist authors; some of his articles are short-form versions of the text included in this new book (cf Senter 2017). His approach is to accept their proposals as valid scientific hypotheses, and not to knock, mock or discount them out of hand from the start. Remember that point; we’ll be coming back to it. This approach means that creationist claims can be considered tested in the empirical sense. It should also be noted that Senter is an Orthodox Christian with qualifications in theology, so his sympathetic and scientifically ‘honest’ approach to creationist claims should not and cannot reasonably be taken as any sort of attack on the Christian faith that the relevant creationists are part of. The fact that Senter is himself religious mean that he can make the argument (should he wish to) that the bad calls and bs put out by creationists is not just ‘bad science’ but ‘bad religion’, too. I’ve heard the same argument from other scientists who maintain an active religious life.

Caption: the book reviewed here is not the first time Senter has written about the ‘fire-breathing dinosaurs’ idea. Image: (c) Skeptical Inquirer.
For completion, and for those who don’t know, I should add that Senter is also an experienced and prolific author of studies devoted to more conventional palaeontological fare: descriptions of new dinosaur species, analyses of phylogenetic patterns, interpretations of functional morphology, and so on. The technical papers of his that I’ve found most useful and interesting include Senter et al. (2004) and Senter (2007) on dinosaur phylogeny, Senter (2006, 2009) on palaeobiology, and Senter (2005), Senter & Robins (2005), Senter & Parrish (2005) and Bonnan & Senter (2007) on dinosaur functional morphology.

Caption: the handsome cover of Senter (2019).
The early chapters of this book evaluate and discuss the creationist contention in general and the relatively young history of the entire movement. The impact of John Whitcomb and Henry Morris’s 1961 book The Genesis Flood is obvious, as is the fact that their arguments fail evaluation (Senter 2019). Nevertheless, their influence was such that – from the early 1970s onwards – a number of like-minded individuals were promoting Whitcomb and Morris’s vision, and were in particular arguing that ancient and medieval writings and works of art make explicit reference to dinosaurs and other long-extinct animals. Senter (2019) uses the term apnotheriopia (meaning ‘dead beast vision’) to describe the tendency of creationist author to interpret monsters in literature and art as long-extinct reptiles.
If apnotheriopia is one of your guiding principles, it ‘follows’ that the fire-breathing dragons canonical to Eurocentric, Christian mythology should be interpreted as dinosaurs or similar reptiles, and that such creatures were dragonesque fire-breathers. So integral has the whole fire-breathing thing been to these authors that they’ve proposed fire-breathing for dinosaurs of several sorts (most frequently hadrosaurs) as well as for pterosaurs and the giant Cretaceous crocodyliform Sarcosuchus (Senter 2019). You might know of one or two cases in which this idea has been mooted. Senter’s book shows that numerous authors have engaged with this vision and written about it. The sheer quantity of this literature is daunting – I was going to say ‘impressive’ but this absolutely seems like the wrong word – and Senter has clearly gone to some considerable trouble to obtain it. He must own a pretty hefty personal library of creationist volumes, and I’m reminded of a statement he makes in one of his papers, wherein he notes that collecting and reading creationist literature on dinosaurs and other extinct animals is one of his “guilty pleasures”.

Caption: some creationist authors have argued that certain dinosaurs could have functioned just like the living bombadier beetles AND SPEWED FIRE!!!!1! One minor issue: bombadier beetles don’t spew fire, they eject hot liquid. Image: Patrick Coin, CC BY-SA 2.5 (original here).
Indeed, the bulk of this book – the long section that runs from chapters 5 through 15 – is a chapter by chapter analysis of the different fire-breathing claims made by creationist authors. These people have, I’ve been surprised to learn, come up with six different mechanisms for fire production in extinct archosaurs. Senter (2019) goes through each in turn, in appropriate detail. In some cases, the proposed mechanisms are total non-starters (no, dear creationists, pterosaurs couldn’t house flammable gases inside their head crests) and can be brushed aside quite swiftly. But in other cases, Senter (2019) has to go down the rabbit-hole of gas chemistry, anatomy and biochemistry, and the history of burns and gaseous explosions in human medicine. All fascinating and well-argued stuff, and full of amazing nuggets of information.

Caption: Parasaurolophus - beloved posterchild of the fire-breathing dinosaurs movement - flames an anachronistic Ceratosaurus, a familiar image from the creationist literature. I believe that this is from one of Ken Ham’s books.
The conclusion, overwhelmingly, is that creationists have been spouting ill-informed (or uninformed) nonsense in coming up with their various fire-breathing fantasies. The proposals concerned are inconsistent with biology, chemistry and physics, and cannot have been present in animals governed by the rules that apply to the living things of planet Earth.

Caption: it’s well known that the crests of lambeosaurine hadrosaurs were hollow, and contained connected internal tubes and chambers. Were these used in the production of fire? No. Image: Sullivan & Williamson (1999).
The book’s final two chapters are connected to the fire-breathing creationist movement, but tackle rather different topics: the origin of dragons as a whole, and the true identity of the biblical Behemoth (Leviathan is covered too), often said by creationists to be a description of a sauropod or similar dinosaur. These two chapters are among the most interesting and valuable in the book.

Caption: why have creationists been so big on the ‘dragons were fire-breathing dinosaurs’ thing? I think it’s partly an effort to attract children to their cult. It isn’t coincidental that most illustrations of fire-breathing dinosaurs appear in books written for children, like this one by Duane Gish.
Even today, the notion that dragons must surely have been based on giant reptiles or reptile-like animals still unknown to science is not unpopular, and is occasionally promoted in the cryptozoological and conspiracy literature. But it’s wrong: the whole idea of dragons as we mostly imagine them (winged, fire-breathing, horned monsters, clad in armour-like scales and equipped with massive limbs and talons) is a mistake, and one that emerged, incrementally, from more mundane origins.
Senter (2019) shows, via statements made in antiquarian literature and by cross-referencing their use of terms, that the term dragon was used – unambiguously, consistently and repeatedly – for snakes, especially for large kinds like pythons. Yes, dragons were snakes. But how does this explain the limbs, wings, fire-breathing and other embellishments? These were added over time, mostly by medieval European authors who were no longer familiar with giant snakes and had heard rumours that dragons could fly (Senter puts this unfamiliarity down to the rise of Christianity and the closing of pagan temples). Feathered wings were added during the 8th century, which then became membranous wings thanks to inventive artists. By the 13th century, dragons were being depicted as quadrupeds (Senter 2019). What about the fire-breathing thing? If dragons were snakes, then some dragons were venomous, and capable of creating a burning sensation in human tissue. Embellish and augment this idea sufficiently, and the concept of fire-breathing winged mega-snakes has emerged. Chinese dragons, by the way, have an entirely independent origin and were mostly based on mammals; Senter (2019) even says that they shouldn’t be called dragons.

Caption: in this most famous depiction of Leviathan - that by Gustave Doré, dating to 1865 - Leviathan is depicted as a monstrous winged serpent of the seas. Image: public domain (original here).
Finally, Senter (2019) also shows – I think convincingly – that the creationist interpretations of both Leviathan and Behemoth of the book of Job are entirely erroneous, but so are the interpretations favoured by the majority of sceptical and ‘mainstream’ authors. I don’t want to steal all of Senter’s thunder, but… Leviathan and Behemoth were both gargantuan mythical serpents, and those authors who have interpreted these creatures as dinosaurs, crocodiles, or big mammals have misunderstood key descriptive phrases, or have been led astray by mistranslations or misinterpretations of the original Hebrew (Senter 2019).

Caption: Senter (2019) uses cartoons like this one to emphasise that Behemoth never was a dinosaur, elephant or hippopotamus, but “a demonic entity that the ancient Hebrews envisioned as a serpent” (Senter 2019, p. 142). The caption to this illustration is “Will be real Behemoth please stand up?”. Image: Senter (2019).
This book is not that lengthy. There are 201 pages, but 45 of them are occupied by a very voluminous bibliography. Plus the book is hardback (or, this edition is, anyway), so appears bulkier than it would do with soft covers. It’s well illustrated and includes numerous colour photos, diagrams of many sorts, and colour cartoons explaining and depicting Senter’s responses to creationist proposals and arguments. There are two things about the images that I dislike. Firstly, some of the colour photos chosen to depict given extinct taxa are quite anachronistic: things would be improved, I feel, if more contemporary reconstructions took their place. Secondly, the colouring used for many of the cartoons is less than great. I mean, the cartoons themselves – which I assume Senter penned himself (he’s a pretty good and competent artist) – are great, but it looks like they’ve been coloured-in with colouring pencils.

Caption: need to feature a depiction of an extinct animal? I, personally, would prefer it if a more up-to-date and aesthetically pleasing image were used in place of this one. Senter (2019) uses several images of models similar to this one when discussing extinct taxa. Image: I’ve been unable to find a source for this picture; it comes from that bottomless pit of hell called pinterest.
So far I’ve been kind to this book. I enjoyed reading it and think it’s a worthy addition to the literature. But I’m afraid that, by the time I’d finished reading it, I’d taken quite a disliking to it, for three reasons.
The first thing I dislike is the way in which creationist claims and proposals are framed. I’m not exactly a fan of creationism, creationist arguments or creationists themselves and I certainly agree with Senter (2019) that the authors who’ve pushed creationist agendas have been scientifically clueless, and/or have sought to wilfully promote anti-scientific gas-lighting. Senter (2019) even finishes the book with a prayer, praising creationist authors for their dogged promotion and energy but wishing and praying that they might make the world a better place by re-directing their energies to something good or constructive. Fair enough.
I do think, however, that Senter (2019) overdoes it in framing creationists and their ideas as ‘silly’ and ‘ridiculous’; Senter (2019) does this throughout the whole of the book such that its entire approach is “let’s all laugh at those whacky creationists” (the subtitle, I’ll remind you, is The Hilarious History of Creationist Pseudoscience at its Silliest). In my opinion (I’d be interested to know if others agree), the book would have worked better if Senter’s approach throughout was neutral and without the mocking. I’ve mocked creationists myself, for sure (Naish 2017), but I’m not about to write an academic book on the subject of their writings. Indeed, given that I’m familiar with Senter’s many papers where he tests creationist claims (all are written in scholarly fashion and use language and phrasing typical for peer-reviewed science), I was surprised to see him follow this path, and I had the impression throughout that it was done in an effort to make the text lighter, more fun, and more appealing. I understand the need for that but I’m saying that – surely – there must have been another way.

Caption: Senter (2019) compares creationist decisions to those made by a character called ‘Silly Chef’ (the muppet-like individual in the middle) who features in a series of cartoons that appear throughout the book. Image: Senter (2019).
Finally on this point, it might be doubtful that the creationists and would-be creationists who are the focus of Senter’s (2019) discussion will ever read this book (it’s abundantly clear that they don’t, or haven’t, read any of the other literature criticising or demolishing their arguments; if they have, they do a good job of making it appear that they haven’t). But by framing the entire book as a “let’s all laugh at those whacky creationists” exercise, the people who might benefit most by reading it will (I assume) be thoroughly put off. Admittedly, this is a moot point anyway in view of my third negative point, but hang on, we’ll get to that in a minute.
The second thing I dislike concerns Senter’s use of humour. This book is well written, and well edited too (I didn’t spot a single typo). But the prose is ruined by Senter’s repeated, very weird forays into simile and metaphor. They are, I’m sorry to say, not just bad, but among the worst examples of writing I can recall. Not only did I not ever find his quips funny, I mostly found them tortuous and daft and I winced every time the text introduced yet another one. I feel bad for saying this and apologise for seeming like a miserable bastard. I disliked this stuff so much that I feel the book would be much improved if all of it was stripped out. And if you’re thinking that – surely – the relevant sections of text can’t be all that bad. Well, here’s one example…
“The misunderstandings and mistranslations necessary to force such an interpretation are almost as bad as those that would be required to infer that the story of David killing Goliath is about a vampire grapefruit preparing a pleasant pile of purple petunias as a fluffy pillow for the happily napping saber-toothed tiger that it keeps as a pet and is convinced for no apparent reason that it is a gigantic German bunny with adorable tiny little ears that wiggle ever so preciously when you gently blow into them” (Senter 2019, p. 144).
There are many other examples of this sort of thing. They ruin the book.
Finally, the third thing I dislike is that great bane of book-buyers from impoverished backgrounds: the price. This book is absurdly expensive; ridiculously so. It’s £69.99 in the UK, $119.95 in the USA (though Amazon is currently selling it at a mere $71.89). As per usual, I appreciate that publishers have to sell books at a given price to cover production costs and to compensate for a sometimes distressingly low number of sales, but I still don’t understand why a slim volume has to cost as much as this one does. Given the price, I fully expected this to be some thick, extremely heavy textbook of perhaps 700 pages or so. But no, it’s a small book of no greater size, production value, academic quality or paper thickness than a great many books half its price or less. I’m extremely pleased to have obtained a review copy but that’s the only way I could ever have obtained it. There is no way I would have purchased it. If a paperback version appears and is reasonably and affordably priced, I apologise for this complaint and may even come back to this review and remove this entire paragraph, but let’s see.

Caption: Senter (2019) is not a big book. Here’s a copy with my hand for scale. Image: Darren Naish.

Caption: I was expecting a much, much larger book. Image: Darren Naish.
I apologise for ending on such a downer.
Fire-Breathing Dinosaurs is certainly a unique book, and – as someone familiar with Senter’s writings on the creationist literature – it does have a magnum opus, end-of-the-road feel about it. As I’ve stated in this review, it’s well-written, has very high production values, and is of significant interest to those who follow the esoteric literature on Mesozoic archosaurs, and on the history of religiously motivated pseudoscience. But it has issues.
Senter, P. J. 2019. Fire-Breathing Dinosaurs? The Hilarious History of Creationist Pseudoscience at Its Silliest. Cambridge Scholars Publishing, Newcastle Upon Tyne. pp. 201. ISBN 978-1-5275-3042-3. Hardback, refs, index. Here at amazon, here at amazon.co.uk, here from the publishers.
A few other reviews of Fire-Breathing Dinosaurs? are online. I deliberately didn’t read them until completing my own review. Having now read them, I see that they make similar points to my own…
Refs - -
Bonnan, M.F. & Senter, P. 2007. Were the basal sauropodomorph dinosaurs Plateosaurus and Massospondylus habitual quadrupeds. Special Papers in Palaeontology 77, 139-155.
Naish, D. 2017. Hunting Monsters: Cryptozoology and the Reality Behind the Myths. Arcturus, London.
Senter, P. 2005. Function in the stunted forelimbs of Mononykus olecranus (Theropoda), a dinosaurian anteater. Paleobiology 31, 373-381.
Senter, P. 2006. Necks for sex: sexual selection as an explanation for sauropod dinosaur neck elongation. Journal of Zoology 271, 45-53.
Senter, P. 2007. A new look at the phylogeny of Coelurosauria (Dinosauria: Theropoda). Journal of Systematic Palaeontology 5, 429-463.
Senter, P. 2009. Voices of the past: a review of Paleozoic and Mesozoic animal sounds. Historical Biology 20, 255-287.
Senter, P. 2017. Fire-breathing dinosaurs? Physics, fossils and functional morphology versus pseudoscience. Skeptical Inquirer 41 (4), 26-33.
Senter, P., Barsbold, R., Brtii, B. B. & Burnham, D. A. 2004. Systematics and evolution of Dromaeosauridae (Dinosauria, Theropoda). Bulletin of the Gunma Museum of Natural History 8, 1-20.
Senter, P. & Parrish, J. M. 2005. Functional analysis of the hands of the theropod dinosaur Chirostenotes pergracilis: evidence for an unusual palaeoecological role. PaleoBios 25, 9-19.
Senter, P. & Robins, J. H. 2005. Range of motion in the forelimb of the theropod dinosaur Acrocanthosaurus atokensis, and implications for predatory behaviour. Journal of Zoology 266, 307-318.
Sullivan, R. M. & Williamson, T. E. 1999. A new skull of Parasaurolophus (Dinosauria: Hadrosauridae) from the Kirtland Formation of New Mexico and a revision of the genus. New Mexico Museum of Natural History & Science Bulletin 15, 1-52.
Mark Witton’s The Palaeoartist’s Handbook
It’s probably – no, surely – true to say that palaeoart (aka paleoart) is more popular right now that it ever has been, a fact due in equal part to a vibrant, active community of people worldwide, to the instant, ubiquitous reach of the internet and the connectedness we feel via social media, to self-publishing and on-demand printing services, and to the excitement and discussion generated by what seems to be a never-ending stream of amazing fossil and anatomical discoveries relevant to ancient animals.

Caption: today, Mark Witton is well known for generating large-scale artworks like this one - depicting the sauropod Diplodocus, and produced to accompanying the NHM’s Dippy specimen as it tours the UK - in addition to work done to accompany press releases. Image: (c) Mark Witton.
It’s no exaggeration to say that Dr Mark P. Witton is, right now, one of the world’s best known and most visible of palaeoartists; his articles and artwork are abundant online, and his work appears in many contemporary published works on prehistoric life, and in various museum installations and other displays. Combine this with the fact that he’s published a long-sought holy grail of the palaeoart canon – a palaeoart handbook – and we surely have one of the most important and worthy palaeoart-themed volumes of all time. Right? Does it deliver?

I refer to 2018’s The Palaeoartist’s Handbook, a slick, extremely affordable softback of 224 pages and extremely high production values. In response to my question above: yes, this book does deliver, and functions extremely well as a ‘handbook’ for those interested in producing palaeoart. Buy it right now if you haven’t done so already. Even those not needing or interested in Dr Witton’s advice should obtain it if they’re interested in palaeoart, since it contains stacks of invaluable review and commentary, does a great job of stating where we are with respect to what we think we know about the appearance of ancient animals, and is really well designed and densely illustrated. It’s probably the most important volume yet published on palaeoart*, and that remains true even if you dislike or disagree with the author’s contentions.
* ‘Importance’ is subjective, but the volume vies – I predict – with 2012’s All Yesterdays and volumes I and II of Dinosaurs Past and Present.

Caption: there aren’t many ‘crucial’/’must have’ volumes on palaeoart, but the Dinosaurs Past and Present volumes are among them, volume II in particular because of Greg Paul’s article (Paul 1987). Images: Natural History Museum of Los Angeles County/University of Washington Press.
A disclaimer I should mention upfront is that Mark and I are long-standing friends and colleagues. I lectured to Mark when he was an undergrad, we’ve been on fieldwork together, and we’ve published several works together on pterosaurs (Witton & Naish 2008, 2015, Dyke et al. 2014, Vremir et al. 2015, Naish & Witton 2017) and palaeoart (Witton et al. 2014). These things might, in theory, mean that I’m positively biased towards his work, but in reality I think they help make me more neutral, since our good relationship means that I can say negative things (where fair and appropriate) and not be worried about being offensive. But let’s see.
Witton (2018) is extremely well designed and very attractive. It’s glossy, full colour throughout, and absolutely packed full of diagrams, photos and art. The art is not just by Mark Witton but also features images by Raven Amos, Rebecca Groom, Johan Egerkrans, Bob Nicholls, John Conway, Emily Willoughby, Julius Csotonyi and Scott Hartman. Holy crap, it’s a virtual who’s who of Early 21st Century palaeoart.

Caption: Witton (2018) includes artwork by several artists, sometimes included to depict diverse styles, compositions and approaches. This is ‘Nemegt Sunrise’ by the amazing Raven Amos (website here), and depicts the oviraptorosaur Conchoraptor with a hermit crab. Image: (c) R. Amos.
What sort of book is this? A ‘palaeoart book’ can be one of several things. It could be a compendium of historical images (like Zoë Lescaze’s gigantic, deeply idiosyncratic but invaluable 2017 Paleoart: Visions of the Prehistoric Past), it could be a bunch of new, daring stuff that makes a point of some sort (cf All Yesterdays), it could be an artist’s porfoilio or a series of portfolios (like the Dinosaur Art volumes, or The Paleoart of Julius Csotonyi), or it could be a technical volume that provides some theoretical or technical background to the field… I’m sure we’re all still waiting for ‘The Grand Handbook to Illustrating Prehistoric Life, a Rigorous How-To Guide’, hint hint.

Caption: the number of books devoted to palaeoart is growing. I think I’ve managed to keep up so far. Image: Darren Naish.
Witton (2018) is partly all of these things: the volume is fundamentally devoted to the techniques, practices and scientific processes and conventions behind the creation of palaeoart, and the case studies and targeted discussions mean that we effectively see much of Witton’s work showcased. But there’s more.
Witton (2018) begins with introductory sections on what palaeoart is and on its history. The historical chapter is quite complete and inclusive, and I’m a big fan of Witton’s take on the work of Cuvier, Hawkins at Crystal Palace and other early efforts. He’s also fair to the artists who produced work during what he terms ‘The Reformation’, some of whom (Greg Paul in particular) have had a major, lasting impact on how we imagine ancient life. The ‘palaeoart meme’ story that I’ve drawn attention to through my own research and the All Yesterdays movement bring a close to this section alongside comments on some of the amazing, exciting new developments being made in the world of soft tissues and palaeo-colour.

Caption: the Crystal Palace animals remain among the most accurate renditions of prehistoric life ever made (like all palaeoartistic reconstructions, they have to be seen as being of their time), and Mark Witton’s take on them is one I absolutely agree with. This photo of the Iguanodon pair was taken in September 2018. Image: Darren Naish.
The meat and potatoes. We then move on to the ‘meat and potatoes’ of the book: a group of chapters that discuss in great detail the process of creating palaeoart. Sections here cover how research is important and how a worker might go about doing it, how knowledge of phylogeny is integral to understanding an organism, and how artists should at least be aware of tropes and stereotypes. This book is fundamentally not a ‘rigorous how-to guide’ to all the prehistoric animals (every time I use this term I’m riffing on the title Greg Paul gave his seminal 1987 article on archosaur reconstruction), but this middle section of the book does include copious discussion of anatomy, the shapes of animals in 3D and cross-section, of musculature and posture, the importance of integument, fat and other external tissue, and so on. I should add that Chapter 9 (‘The Life Appearance of Some Fossil Animal Groups) is devoted to the probable life appearances of key tetrapod groups. Ha, take that fishes.

Caption: in many cases, the favoured, traditional look for a given prehistoric animal is not necessarily the one we might favour. Here’s an example: Mark has argued that a new look for the proboscidean Deinotherium - shown here - should be considered. Image: (c) Mark Witton/Witton (2018).
These central chapters are probably the most important part of the book and will be those used most by the largest number of people. There’s a ton of information and discussion, and Mark describes in detail how he’s arrived at the conclusions he has. Readers of Mark’s blog will be familiar with some of the arguments here and might also know that much of it has never been properly published (as in, in technical papers or articles). The book is therefore especially significant as a source of primary data, though I know that efforts are underway to get at least some of it into the primary literature.

Caption: extensive sections of Witton (2018) discuss osteological correlates for external texture and other features. In some cases - like ceratopsian dinosaurs - there are many such correlates. Image: (c) Mark Witton/Witton (2018).
How confident can we be that Mark is ‘right’ when it comes to his arguments about lips, cornified facial tissue, scalation or fuzziness on the body and so on? I think that a strong response would be that he has at least described, explained and illustrated his reasoning and it’s difficult to think that he’s ‘wrong’, two caveats being that there is – as Mark states quite clearly – still some considerable slop as goes determining the relative size of keratinous coverings (like the pads, scales and sheaths covering horns, claws and so on), and that the vagaries of taphonomy might still be cheating us out of valuable information on which archosaurs had filaments, fuzz or feathers. Yes, I still think that big tyrannosaurs could have been fuzzy and that we aren’t picking this up because the fossils concerned aren’t preserved in the ideal sedimentological regimes.
Speculation and the All Yesterdays Movement. The main message here is that while some speculation always has to be included in palaeoartistic reconstructions, there’s a lot of stuff that’s knowable, or potentially knowable, and informed by actual anatomical data. This is increasingly the case even for colour and pattern (caveat: we still only have data on some infinitesimally tiny percentage of extinct animals). The door is not open for any possibility, and artists who wish to be seen as doing work that’s scientifically credible have to take into account data derived from fossils as well as ‘rules’ (or guidelines) gleaned from living animals. However…

Caption: Conway et al.’s 2012 All Yesterdays has changed the way many people approach palaeoart… but is this for better, or for worse? Image: Conway et al. (2012).
A valid, controversial and timely point concerns just how much speculation is permissible in palaeoart. This is something I feel especially connected to given the impact of my 2012 book – co-authored with John Conway and C. M. Kosemen – All Yesterdays (Conway et al. 2012) and the subsequent ‘All Yesterdays Movement’, which is hated by some but loved by others. Mark’s take on what happened post-AY is that a lot of AY-inspired artwork has failed to appreciate the nuance of the original work, and that AY was (wrongly) taken by some as a green light to go nuts and do whatever, the results being misguided and likely wrong.

Caption: Mark has indulged in some speculation himself (here: shaggy-coated pachyrhinosaurs), and it’s down to opinion as to whether this is as extreme as anything depicted in All Yesterdays. Image: Mark Witton/Witton (2018).

Caption: it would be wrong to avoid bringing attention to Franco Tempesta’s woolly, cold-adapted pachyrhinosaur, very obviously inspired by Mark Witton’s, and appearing in the 2016 Usborne book Build Your Own DInosaurs Sticker Book. I was consultant, but I’m sure that that’s coincidental. Image: (c) Franco Tempesta/Usborne.
I agree… if we’re talking about artworks that aim to reflect possible realities. A nuance to the nuance of AY is that there exists a small contradiction in the aims of its creators. Yes, we argued that there are many potentially valid, scientifically defensible possibilities that hadn’t or haven’t been sufficiently explored in pre-AY palaeoart, but we did also promote the idea that people might explore other possibilities – even those weird or dumb or wrong – purely for the sake of artistic expression. That this view is canonical in the AYverse is demonstrated by the inclusion in the sequential All Your Yesterdays of retrosaurs that are absolutely contradicted by data but still fun from an artistic take. In other words, not all AY-inspired art is meant to be scientifically defensible. The takehome – post-AY – is that people need to say what they’re aiming to depict: a random fancy or a serious proposal?

Caption: not all AY-inspired art is meant to be scientifically responsible and potentially realistic, some of it is deliberately whimsical and fanciful. Exhibit A: the wonder that is Spinofaaras vulgaris, a creature that now has an internet life of its own. Image: (c) Chris Masna (original here).
Anyway, returning to the contents of Witton (2018), this main section wraps up with some thoughts on how landscapes are created, and how composition and mood can be formed. Discussing environments and landscapes involves science – geology, geomorphology and palaeoclimate, among other things – and this is Witton’s main strength, but I also found his take on composition, stylistic choices and other matters of artistic style compelling…. speaking as someone who lacks artistic training and expertise, that is. The volume ends with a chapter on the professional side of palaeoart.
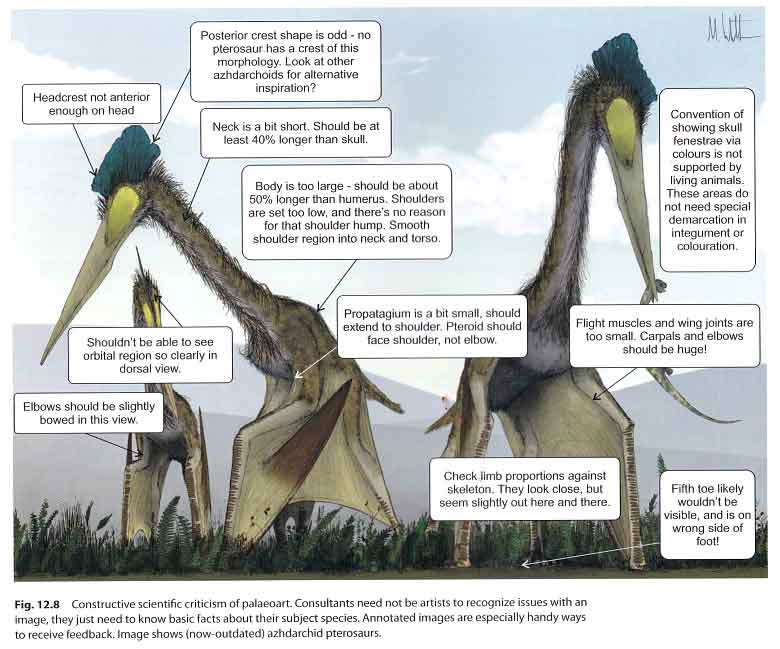
Caption: feedback and criticism is crucial, but it can be difficult to know what to say to artists when you aren’t one yourself. In this section of the book, Mark provides advice, using his 2008 azhdarchid image as a piece that might benefit from constructive criticism. This piece accompanied the Witton & Naish (2008) PLoS paper on azhdarchids. Image: (c) Mark Witton/Witton (2018).
On the negative side of things… I find the editing sloppy in places and think that the prose could have been tightened here and there. There are also a few turns of phrase that I found awkward, weird or (sorry) terrible, top of the list being the reference to “palaeontologists with the mightiest beards” (p. 38). On technical aspects, I’m a bit confused by Mark’s use of ‘reptile’ in the old, paraphyletic sense (in a volume otherwise using modern phylogenetic nomenclature, wouldn’t it make sense to use Reptilia for the lizard + turtle + croc clade, and not to use it for a paraphyletic assemblage that excludes birds?). Panoplosaurus is wrongly called an ankylosaurid (p. 125), and isn’t Deinotherium a deinotheriid, not a deinotherid? These are minor, piffling, trivial things that I only state here because I have nowhere else to put them.

Caption: the book is just full of spectacular imagery like this, much of which hasn’t appeared in print before. This image depicts the azhdarchoid pterosaur Thalassodromeus. Image: (c) Mark Witton/Witton (2018).
All in all, The Palaeoartist’s Handbook is an excellent, beautifully produced, well crafted book which contains a wealth of information on the life appearance of extinct animals and how we might imagine them as living things, and it’s phenomenally good on the workings of palaeoart more generally. It should have broader appeal than to the palaeoart fraternity alone, and I think that anyone seriously interested in prehistoric animals or even in the history of art or the way people have imagined the past should obtain it too. For now, Witton (2018) is – mission fulfilled – THE palaeoartist’s handbook indeed.
Mark P. Witton. 2018. The Palaeoartist’s Handbook: Recreating Prehistoric Animals in Art. The Crowood Press, Marlborough (UK), 224 pp, softback, index, refs, ISBN 978-1-78500-461-2. Here on amazon. Here on amazon.co.uk.
For previous TetZoo articles on palaeoart and Wittoniana (the ver 2 and ver 3 ones have been ruined by removal of images), see…
The Great Dinosaur Art Event of 2012, November 2012 (but all images now missing)
All Yesterdays: Unique and Speculative Views of Dinosaurs and Other Prehistoric Animals - the book and the launch event, December 2012 (but all images now missing)
Artistic Depictions of Dinosaurs Have Undergone Two Revolutions, September 2014 (but all images now missing)
The Dinosaurs of Crystal Palace: Among the Most Accurate Renditions of Prehistoric Life Ever Made, August 2016
The Natural History Museum at South Kensington, November 2016
Naish and Barrett's Dinosaurs: How They Lived and Evolved, November 2016
25 Years after Jurassic Park, Part 1, June 2018
25 Years after Jurassic Park, Part 2, June 2018
25 Years after Jurassic Park, Part 3, June 2018
Dinosaurs in the Wild: An Inside View, July 2018
Recollections of Dinosaurs Past and Present, the 1980s Exhibition, February 2019
Refs - -
Paul, G. S. 1987. The science and art of restoring the life appearance of dinosaurs and their relatives - a rigorous how-to guide. In Czerkas, S. J. & Olson, E. C. (eds) Dinosaurs Past and Present Vol. II. Natural History Museum of Los Angeles County/University of Washington Press (Seattle and London), pp. 4-49.
The Fate of Burian’s Styracosaurus
Among the most recognisable staples of popular prehistoric animal books is the multi-spiked North American ceratopsian dinosaur Styracosaurus albertensis, discovered in Alberta in 1913 and described and named later that same year by Lawrence Lambe.

One of my several memorable childhood encounters with Styracosaurus was in the 1975 movie The Land That Time Forgot, a World War I adventure film based on a 1918 novel by Edgar Rice Burroughs. If you haven’t seen The Land That Time Forgot, it revolves around the discovery of a lost land called Caprona by the crew of a German U-boat. The main cast are not all German, since they’ve taken aboard a bunch of British people and even an American, all rescued from their own sinking merchant vessel. Doug McClure is the main star.

Caption: screengrab from The Land That Time Forgot, showing the styracosaur that doesn’t get shot.
At least some of my childhood takes on prehistoric animals and their world were inspired by that film, and one scene I remember in particular is a night-time segment in which two unlucky styracosaurs are fired upon by the U-boat. One is hit (one of its characteristic frill spikes is blasted off) and dies, a symbolic tear trickling from its eye. It was thus a great thrill for me to recently see this model, at the 2019 Portsmouth Comic Con. Yes, it’s the intact one of the two The Land That Time Forgot styracosaurs.

Caption: Roger Dicken’s Styracosaurus, as seen at Portsmouth Comic Con in May 2019. Note the massively wide, deep snout and prominent depressions on the frill. Image: Darren Naish.
Why was this model at Portsmouth Comic Con? Because movie model-maker Roger Dicken was there, and I got to speak to him. Roger’s IMDB page gives some idea of how many movies he’s been involved in during his long and illustrious career: he made the original Alien chestburster, among many other things. In speaking with him, I was finally able to confirm something I’d always suspected: the styracosaurs in the movie were based very specifically on the ones illustrated by famous Czech palaeoartist Zdeněk Burian (1905-1981) for his grand 1972 book with Zdeněk V. Špinar, Life Before Man (Spinar 1972). Burian illustrated Styracosaurus several times during his career, but this painting (actually produced in the 1940s, not the 70s) is the most familiar and most reproduced. The animal is broad across the muzzle, has distinct sunken regions on the frill, and the spikes on the frill (in the background individual) closely follow the contours of the shoulder and back.

Caption: one of so many beautiful and hugely influential scenes of ancient life by Zdeněk Burian. High-quality reproductions of this image reveal far brighter colours than those normally obvious in books (like a red ring around the eyes), and extra details like a scaly fringe to the side of the upper beak. Though best known for versions published in the 1970s, it was produced in 1941. Image: (c) Zdeněk Burian.
Burian’s art was – and arguably still is – highly influential, not just because it’s wonderful and looks amazing but also because it was just about the only palaeoart accessible to a large sector of the interested public during the 1960s and 70s. It’s no surprise that the look he favoured for a given animal often became the standard template for the species concerned. But how did Burian himself work out what ancient organisms looked like? He was working at a time when information was scant, experts were few and hard to communicate with, and literature non-existent or highly technical. We know that Burian consulted extensively with Špinar, and also that he used measurements and images of fossils to inform the reconstruction process.
But…

As recently realised and brought to my attention by Mark Witton, it turns out that Burian’s take on Styracosaurus wasn’t exactly unique. Look at this image (above) by Vernon Edwards, apparently made during the 1930s. Edwards made a huge number of dioramas depicting prehistoric animals in landscapes, many of which are depicted in books of the early 20th century (my main source for these images being the 1941 The Miracle of Life, which I’ve written about before [images now removed from article, well done SciAm]). The similarities between the Burian and Edwards scenes are many. The composition and landscape is similar, the animals are posed the same way as goes the angles we see, and there are lots of anatomical similarities. All those features I mentioned above are visible, and note also the bulging neck creases visible on the animal we see in profile.

Caption: see that dated signature at bottom? The date is ‘41’, and if you’re not convinced, look below…

Caption: a detail of Burian’s best known Stegosaurus painting, again showing ‘41’. Image: (c) Zdeněk Burian.
So - is this a case of Burian basing his work on that of a previous artist? As noted above, Burian’s styracosaur scene is from 1941 (the date is obvious in good versions of the image; see above). Edwards’s scene is supposedly from the 1930s, but the oldest published version I’ve seen is from 1941. Could it be, then, that Vernon Edwards produced this image in 1941 – not during the 30s – and that it was based on Burian’s scene, not vice versa? I honestly don’t know and haven’t been successful in working out the exact details on what happened.
If Burian did base his work on the image by Edwards, this might be – as Mark stressed in a twitter exchange – the only case in which Burian based his work on that of another palaeoartist. It’s not as if we’re saying that he was a regular plagiariser or anything.

Caption: at left, the two Ladybird books discussed below. My copy of the 1974 Dinosaurs has a bright pink scribble across its cover. At right, a bonus Burianesque styracosaur depicted on the cover of another Ladybird book. Images: Darren Naish, Arran Alexander Collection.
As mentioned earlier, Burian’s work was so influential that it was widely used by other artists. At this point I could write a great deal about Burian-inspired images of this dinosaur, but I’ll finish by discussing one in particular. I don’t know how familiar Ladybird books are outside of the UK (non-UK readers, let me know), but – in the UK – they’re among the most beloved and cherished of books to people who grew up between the 1950s and 90s. My favourite was always, and still is, the 1974 Labybird leader book Dinosaurs, authored by Colin Douglas and illustrated by Bernard Robinson (Douglas 1974). And there on page 38 we find this striking image, featuring a stormy sky and a totally anachronistic Tyrannosaurus (Styracosaurus is some millions of years older than Tyrannosaurus)…

Caption: the Ladybird/Bernard Robinson vision of Styracosaurus.
Such was the popularity of this book that an enlarged and augmented edition – titled Dinosaurs and Prehistoric Animals – appeared in 1978, with extra illustrations and much more text (Wellfare 1978). It enabled the art to be shown at larger size but features many of them in cropped form such that their relationship to larger scenes is unfortunately ruined. Anyway, here’s the spectacular styracosaur again. It has a fantastic eagle-like glint and hint of simmering rage in its eye. The spines around the edge of the frill look to be based on Burian’s painting more than on an actual styracosaur fossil, and the scaly edge to the beak - again, inspired by what Burian depicted - is an interesting touch since it shows that the artist was seemingly unaware of the presence of keratinous beak tissue in these animals (a thing they surely had).

Caption: Styracosaurus from the 1978 Dinosaurs and Prehistoric Animals. Image: Ladybird/Bernard Robinson.
How has the Burian-esque view of Styracosaurus fared in more recent decades? Our improved understanding of ceratopsian musculature and skin texture – combined with our rather more dynamic view of what Mesozoic dinosaurs were like overall – means that any good modern take on Styracosaurus shows a more active beast with more erect limb carriage and more elevated head and neck. The snout shouldn’t be massively wide and turtle-like as Burian (and Edwards) showed, but narrower and deeper, and it should also be more obvious that the spikes around the edges of the frill are distinct, independent structures, not outgrowth of the frill’s main body.
I included a section on Styracosaurus in my 2009 book on the history of our building knowledge on dinosaurs, The Great Dinosaur Discoveries (Naish 2009). It’s a decent potted history of what we know of Styracosaurus, culminating with the revision and redescription of the styracosaurs published by Ryan et al. (2007). Ryan et al. (2007) recognised two Styracosaurus species but the second of these – S. ovatus, named in 1930 – is currently regarded as belonging to the distinct genus Rubeosaurus.

Caption: the Styracosaurus skeleton AMNH 7372, originally named as the distinct species S. parksi in 1937 (but now regarded as synonymous with S. albertensis). Image: AMNH/public domain.
Little known away from the ceratopsian research community is that the lower jaw and skeleton of Styracosaurus wasn’t collected from the field until 1935 (remember: this dinosaur was named and described in 1913). The nasal horn of the original skull was broken. Lambe thought that this break had occurred half-way along the horn’s length and reconstructed the missing tip accordingly, the result being a ceratopsian with a very long and straight nasal horn perhaps 60 cm long. More recently discovered specimens show that his assumption – while sensible – was incorrect, and that the horn was actually shorter and blunter than he’d concluded, and that 30 cm would be a more realistic length (Ryan et al. 2007) (caveat: I’m talking here about the bony core of the horn, not the keratinous covering). Old reconstructions therefore exaggerate the length of that horn.

Caption: the skull of Styracosaurus albertensis, as seen from the front. The beak section is narrow and deep, not wide and rounded. This is AMNH 7372 in New York, collected from what’s now Dinosaur Provincial Park by Barnum Brown in 1915. Image: Claire Houck, CC BY-SA 2.0 (original here).
That’s where we’ll end things for now. There’s tons more to say about how ceratopsians have been depicted in life and on what we think we know about their anatomy and biology. I’ve written a lot about these issues in the past, but nearly everything has been ruined due to the removal of images at ScienceBlogs and SciAm.
For other TetZoo articles on ceratopsians and related palaeoart-themed issues, see…
A very alternative view of horned dinosaur anatomy, April 2009 (but now lacking all images)
Dinosaurs and their exaggerated structures : species recognition aids, or sexual display devices?, April 2013 (but now lacking all images)
Palaeoart Memes and the Unspoken Status Quo in Palaeontological Popularisation, February 2017 (paywalled for me as I’ve used up my SciAm quota)
The Ridiculous Nasal Anatomy of Giant Horned Dinosaurs, November 2016
Refs - -
Douglas, C. 1974. Dinosaurs. Ladybird Books, Loughborough.
Naish, D. 2009. The Great Dinosaur Discoveries. A&C Black, London.
Ryan, M. J. Holmes, R. & Russell, A. P. 2007. A revision of the late Campanian centrosaurine ceratopsid genus Styracosaurus from the Western Interior of North America. Journal of Vertebrate Paleontology 27, 944-962.
Špinar, Z. V. 1972. Life Before Man. Thames and Hudson, London.
Wellfare, G. 1978. Dinosaurs and Prehistoric Animals. Ladybird Books, Loughborough.
A postscript…
I’ll just leave this here. The image at top is (c) Robert Bakker, and was produced in 1971. The image below it is by Burian and is dated 1976. Montage from here at Earthling Nature.

Recollections of Dinosaurs Past and Present, the 1980s Exhibition
As a regular reader here, you should be familiar with my interest in the portrayal of dinosaur life appearance, and indeed in palaeoart in general. It’s now trite to explain how the iconic, well-drafted, anatomically rigorous illustrations of Robert Bakker, Greg Paul, Mark Hallett and others were integral to the popularisation and dissemination of the ‘dinosaur renaissance’ that occurred between the late 1960s and 1990s, and few interested in prehistoric life will have failed to notice how quickly and frequently reconstructions of extinct species – often good, accurate and innovative – appear today, typically in the digital medium. Palaeoart remains relevant, essential, and with a huge fanbase.

Caption: Robert Bakker’s 1969 sprinting Deinonychus, produced to accompany John Ostrom’s seminal article on this amazing dinosaur. Artistic depictions like this one cement the idea that art has conveyed scientific concepts to the public... but you’ve heard all that before. This is one of several Bakker images included within the exhibition discussed in this article. Image: (c) Robert Bakker.
During the late 1980s and early 90s, a remarkable thing happened. A team at the Natural History Museum of Los Angeles County, assisted by funding from the Natural History Museum Foundation and fronted by guest curator Sylvia J. Czerkas, created a travelling palaeoart exhibition titled Dinosaurs Past and Present. Opening in Los Angeles in February 1986, and accompanied by a symposium that featured talks on dinosaur science and artwork, the exhibition featured a selection of historical illustrations, paintings and sculptures as well as the very best of contemporary work.

Caption: the entrance to what might have been the best palaeoart-themed exhibition of all time, as seen at London’s Natural History Museum in late 1990 and early 1991. A better quality version of this photo will be uploaded to TetZoo in the near future (thanks to Spike Ekins for permission). Image: Spike Ekins.
Accompanying literature does little to shed light on the backstory of the project and how it all came together – maybe this information is out there and I just haven’t seen it – but the final exhibition essentially functioned as a ‘who’s who’ of late 1980s palaeoart. Credits and acknowledgements show that work was loaned from museums and publishers, but overwhelmingly from private collections, mostly those of the artists themselves. This is interesting for several reasons, one being that the curators and organisers clearly had good relations with the artists, another being that the vast majority of iconic 1980s palaeoart was – as of 1986 at least – owned by its creators and not by the sort of wealthy socialites often associated with niche art. Alas poor palaeoart and its lack of reliable patrons (can someone get that printed on a t-shirt please).
After its Los Angeles opening in 1986, the exhibition toured North America, stops including the Smithsonian, the AMNH, the Tyrrell and the Field. It then crossed the Atlantic for showings in Edinburgh, Cardiff and London, where it finished its run in the January of 1991. Sorry, non-English-speaking nations.

Caption: London’s Natural History Museum is an amazing building, decorated inside and out with images of plants and animals. The pterosaur at left is one of several visible on the outside of the museum. Images: Darren Naish.
I didn’t visit London’s Natural History Museum all that often during my teenage years, but I did get to go there several times as special birthday trips. It was on one of these that – in 1990 – I got to see Dinosaurs Past and Present. This was a total accident, by the way, and not due to clever planning. I didn’t even know about the exhibition beforehand, or that it would be on show during my visit.
Two accompanying volumes – edited by Sylvia J. Czerkas and Everett C. Olson – were produced for the exhibition (Czerkas & Olson 1987a, b). They’re must-haves for serious students of palaeoart, containing many articles that provide invaluable background, discussion and commentary. Those by Mark Hallett, Greg Paul and Dale Russell are especially good. They also, it has to be said, contain several articles that are less valuable, are not especially relevant to palaeoart, and could well have appeared elsewhere.

Caption: the covers of volumes I and II of Dinosaurs Past and Present. Both were initially published (in 1987) in hardback, and later (1989) released as softback. My copies are softbacks. Images: Natural History Museum of Los Angeles County/University of Washington Press.
An introduction by John M. Harris (1987) provides some background to the exhibition and its accompanying symposium. Harris’s article, incidentally, includes that specific statement where ‘reconstruction’ and ‘restoration’ are explained to mean different things (p. 4), so it might be this article – rather than the one by Mark Hallett in the same volume – that is best cited as the origin of this concept.
As one of the few people who (a) is of the right age and (b) writes regularly about palaeoart, I’ve been feeling increasingly duty-bound to discuss my recollections of seeing Dinosaurs Past and Present in London in 1990. What, then, do I remember?
First things first: photos were not allowed. You might like to decide among yourselves whether this is a good thing or not. So, despite having a camera with me (a Canon 35 mm of some sort, I think) I didn’t take any photos, except a few sneaky, illegal shots of Stephen Czerkas’s Allosaurus model. More on that model below.

Caption: here’s the only photo I’ve seen that shows what the exhibition looked like. Behind the Czerkas Allosaurus, framed illustrations on the wall include pieces by Ken Carpenter (at left) and Greg Paul (at right). I can definitely see Paul’s Styracosaurus vs Albertosaurus image. A better quality version of this photo will be uploaded to TetZoo in the near future (thanks to Spike Ekins for permission). Image: Spike Ekins.
As goes the exhibition as a whole, I recall it being in one of the galleries that are perpendicular to the marine reptiles corridor (though I could be wrong). The illustrations were hung on white panelling erected to cover the walls (as confirmed by the photos here, by Spike Ekins), there also being a white partitioning wall, reaching to chest- or shoulder-height, erected along the middle of the gallery. Illustrations were hung from either side of this partition as well. There must have been a shop somewhere, since I distinctly recall the two volumes of Dinosaurs Past and Present being on sale. I was able to buy one of them during my visit (I went for Volume II), eventually obtaining Volume I some years later on a separate trip. I recall these books as being cripplingly expensive and my buying of Volume I consuming essentially all of my Natural History Museum trip budget, so it’s embarrassing now to see how (relatively) inexpensive they actually were.

Caption: my copies of the Dinosaurs Past and Present books still contain their original price labels. Oh, not as expensive as I remembered, then. Whatever. Image: Darren Naish.
Volume I of Dinosaurs Past and Present includes a checklist of all the art featured in the exhibition. No less than 144 pieces were included, the majority being paintings and drawings. They were arranged chronologically, pieces by the likes of Benjamin Waterhouse Hawkins, Charles Knight and Richard Swan Lull being nearest the entrance and the more contemporary work of Mark Hallett, Ely Kish, William Stout, Greg Paul and so on being encountered later on.

Caption: Charles Knight’s Dryptosaurus painting of 1897 was included in the exhibition and was loaned from the American Museum of Natural History. It’s 58 cm long, 40 cm tall. Image: public domain, wikipedia (original here).
Among the older works, I recall seeing the 1879/1880 ‘Pleasures of Science’ piece by Arthur Lakes, and the Benjamin Waterhouse Hawkins depictions of Iguanodon, Hylaeosaurus and Megalosaurus. Weirdly, I have no firm recollection of seeing the works of Charles Knight, Rudolph Zallinger or Peter Zallinger, which is distressing given how influential they’ve been.

Caption: Rudolph Zallinger’s incredible mural at Yale’s Peabody Museum is virtually never shown in published photos, and most of the images you’ve seen - like this one - are from the prototype ‘Study’, in which the animals look quite different. Image: (c) Rudolph Zallinger/Peabody Museum of Natural History, Yale University.
In fact, most of what I remember is decidedly contemporary. Of the Mark Hallett originals on show, I remember the montage images that show representative members of assorted clades. My strongest memory is of the ornithopod group, partly because I recall noticing that a hadrosaur originally meant to be a Tsintaosaurus (and shown with the classic, erroneous ‘unicorn’ crest*) had been ‘corrected’ such that it was now an Edmontosaurus. Hallett’s ‘Iguanodon studies’ is also allotted to memory (perhaps because I always liked his idea that iguanodontian beak tissue might have a serrated, pseudotoothed look), as well as his large Morrison Formation panorama of 1975.
* Tsintaosaurus, it turns out, was neither unicorn-crested, nor flat-headed (as has been argued), but instead equipped with a tall, bulbous crest (Prieto-Márquez & Wagner 2013). No more phallic jokes then.

Caption: Mark Hallett’s ‘Crossing the Flats’ was at the exhibition. It’s a big piece, more than 1.2 m long. Today we don’t think that Mamenchisaurus looked quite like this (Mark was basing his reconstruction on the idea that Mamenchisaurus was a diplodocid). This painting always interested me because of the bipedal, narrow-chord pterosaurs as much as the sauropods. Image: (c) Mark Hallett.
I also recall Ken Carpenter’s illustrations of Sauropelta and ‘Velociraptor’ (proof that Ken was following Greg Paul’s nomenclature; the ‘Velociraptor’ here actually being Deinonychus).
The Greg Paul Years. I make no secret of the fact – it’s mentioned in most books and articles I’ve written or contributed to on dinosaurs and palaeoart (Naish 2009, Conway et al. 2012, Naish & Barrett 2018) – that I consider Greg Paul one of the most influential and important of palaeoartists, and this is true even if you disagree with various of his contentions. Indeed, Greg’s significance in palaeoart is demonstrated by the fact that a full 24 of his pieces were included within the exhibition, more than any other artist.

Caption: Greg Paul’s 1987 article from Dinosaurs Past and Present, Volume II remains one of the best guides to the life appearance of extinct archosaurs, even though it’s now substantially dated. John Conway and I aim to produce a volume that ‘replaces’ it at some point; meanwhile there’s Mark Witton’s excellent The Palaeoartist’s Handbook (which will be reviewed here at TetZoo soon). Image: Darren Naish.
Unsurprisingly, then, my main recollections concern his pieces. His black and white artwork is mostly small (say, 40 x 40 cm or so), numerous corrections and edits being visible where they were made either with paper whitener (we tend to call it tipex in the UK due to one specific brand) or with pieces of card that have been stuck over the relevant areas. I distinctly recall the tail of one of the tyrannosaurs in his ‘Monoclonius albertensis Fending Off Albertosaurus libratus’ (“Monoclonius albertensis” = Styracosaurus) revealing obvious signs of having its angle corrected (a familiar issue to those who’ve drawn dinosaurs). His ‘What Happens When Apatosaurus ajax Seeks Aquatic Refuge from Allosaurus fragilis’ originally had the riverbed too high, a correctional piece of card allowing him to position it further down the canvas, thereby allowing more space for the swimming sauropod’s feet.
I’m pretty sure I also recall looking at his iconic Giraffitan scene, at his G. brancai muscle study, and at the scene showing Iguanodon and Mantellisaurus (I. atherfieldensis at the time) foraging alongside one another.

Caption: Greg Paul’s art is among the most influential dinosaur-themed palaeoart ever produced. Today we know that his feathered non-bird theropods aren’t feathery enough but, hey, you have to start somewhere. This painting was featured in the exhibition. Image (c) Greg Paul.
I also recall a few colour Greg Paul paintings. I definitely remember the ‘Resting Velociraptor antirrhopus Pair’ painting, perhaps in part because Greg’s ideas on feathered dinosaurs, and his taxonomic argument that Deinonychus should be considered a species of Velociraptor (which he’s since abandoned), were novel to me at the time. I think that the Allosaurus vs Diplodocus scene was there but my recollection is hazy. I’m far more confident about seeing Paul’s painting of the Pentaceratops herd. I remember it being pretty big, a metre or so in width. To, again, my disappointment, I have no recollection whatsoever of seeing the ‘Tyrannosaurus torosus in a Fast Run’ (for “Tyrannosaurus torosus” read Daspletosaurus). Shocking, because this is another iconic image of the Dinosaur Renaissance.

Caption: Greg Paul’s running Daspletosaurus - here on the cover of Lauber (1989) - is one of his most famous paintings, yet I don’t remember seeing it. Oh well. Image: Darren Naish.
Enough with the Greg Paul. What else do I remember? I do distinctly remember seeing colour pieces by John Gurche. Gurche has the most incredible style, his colour pieces looking like photos and very often including a marked contrast between a brightly lit, extremely sunny portion and a pitch-black area of deep shadow.
Some of Gurche’s paintings are surprisingly small. On seeing the 1982 Archaeopteryx piece, I was struck by its small size (around 20 x 30 cm). I don’t remember seeing his 1985 Daspletosaurus vs Styracosaurus piece, but it was definitely there. Again, what is wrong with my memory?

Caption: a John Gurche painting of 1985 appeared on the cover of an especially famous and influential book (Bakker 1986). Image: Penguin Books.
On that note: bizarrely, I don’t remember seeing pieces by Doug Henderson, Robert Bakker or Ely Kish, even though they were definitely there too. Weird. Swiss cheese memory. Nor do I remember seeing the original egg tempera study of Zallinger’s Yale mural, Hallett’s ‘Crossing the Flats’, or a hundred other significant pieces that were there. The more I think about this, the angrier I become. Maybe all that drinking and recreational drug use is to blame. Ha ha, kidding, kidding, kidding.

Caption: for its stint at the Royal Museum of Scotland in Edinburgh, publicity for Dinosaurs Past and Present included this photo-shoot, involving the late palaeontologist Beverly Halstead and Ron Seguin’s troodontid and dinosauroid models. Halstead is at far right. I don’t know if Halstead is goofing around (he has his arm around the dinosauroid’s shoulders), but it looks like he is. Image: (c) New Scientist.
Finally, what about models? Yes, the dinosauroid and the accompanying Stenonychosaurus were both there, and oh does it hurt to not have any photos of them. You’ve surely seen colour photos of both of these models (created by Ron Seguin in co-operation with Dr Dale Russell): they’re reproduced in many books. Large potted plants (including parlour palms and cycads) were arranged around the models to create a slightly greener vibe. You can see all of this in the black and white photo reproduced above, taken to show the late Beverly Halstead with the models while Dinosaurs Past and Present was on show in Edinburgh.

Caption: one of several books that were highly inspirational if you encountered them at the right time. Wallace (1989) includes both exciting artwork as well as cutting-edge news (Protoavis?! Wtf!, I thought). And on the cover? Yes, it’s Czerkas’s allosaur, portrayed as a dark and sinister predator: “the Darth Vader of animals”, to quote John Conway. Image: Darren Naish.
Nearby, Stephen Czerkas’s half-life-size Allosaurus model was on show. As mentioned above, this is the one exhibit I have photos of (though I can only find one of my photos today, dammit). At half life-size, it stands perhaps 1.5 m at the top of the head. It was browner than I always imagined: based on the cover of Joseph Wallace’s 1989 The Rise and Fall of the Dinosaur* – an influential book for the young Darren Naish – I always thought it was dark grey or black. Like all of Czerkas’s models, it included a spectacular and really impressive amount of detail and was a joy to look at.
* I find it a bit annoying that Steve Brusatte’s new book has essentially the same title. Steve didn’t know of Wallace’s book until I told him about it, so not his fault. Did no-one think to say anything?

Caption: Stephen Czerkas’s brilliant Allosaurus model at the Natural History Museum, London (as demonstrated by the accompanying stonework) in 1990. Note that the model is in a different position relative to where it is in Spike’s photos shown above. Image: Darren Naish.
Several smaller models, made by Stephen and his wife Sylvia Czerkas, were also on show and on top of white, rectangular display stands. I remember Sylvia’s hatching Protoceratops and, I think, Stephen’s 1986 Stegosaurus, controversially constructed with the single row of plates that Stephen thought correct (Czerkas 1987).
For completist reasons I should note that I did get to meet Stephen, once, at the Denver 1999 Society of Vertebrate Paleontology meeting. We spoke briefly about ‘Archaeoraptor’, later unveiled as a composite but thought by Czerkas at the time to be a long-tailed early bird, but I never did get to talk to him about his art.
Anyway, that about wraps things up. Clearly, my memory of a truly momentous and significant exhibition is alarmingly deficient, and the fact that I don’t really have any photographic record of what I saw just makes it worse. What I do remember was, however, thrilling. There have, of course, been a good number of palaeoart-themed exhibitions since 1991, but I think it’s fair to say that none have been as grand, culturally significant or momentous as Dinosaurs Past and Present. Could its like ever occur again? Maybe time will tell, or maybe I’m nostalgic for a Golden Age which has long since passed.

Caption: what has happened in the world of palaeoart since the days of Dinosaurs Past and Present? Quite a lot… Image: Darren Naish.
Palaeoart and changing ideas on the life appearance of Mesozoic dinosaurs have been covered at TetZoo many times over the years. Many of the older articles are now useless because malfunction at the hosting sites has removed their illustrations. Anyway, see…
The Great Dinosaur Art Event of 2012, November 2012 (but all images now missing)
All Yesterdays: Unique and Speculative Views of Dinosaurs and Other Prehistoric Animals - the book and the launch event, December 2012 (but all images now missing)
Artistic Depictions of Dinosaurs Have Undergone Two Revolutions, September 2014 (but all images now missing)
The Dinosaurs of Crystal Palace: Among the Most Accurate Renditions of Prehistoric Life Ever Made, August 2016
The Natural History Museum at South Kensington, November 2016
Naish and Barrett's Dinosaurs: How They Lived and Evolved, November 2016
25 Years after Jurassic Park, Part 1, June 2018
25 Years after Jurassic Park, Part 2, June 2018
25 Years after Jurassic Park, Part 3, June 2018
Dinosaurs in the Wild: An Inside View, July 2018
Refs - -
Bakker, R. T. 1986. The Dinosaur Heresies. Penguin Books, London.
Czerkas, S. A. 1987. A reevaluation of the plate arrangement on Stegosaurus stenops. In Czerkas, S. J. & Olson, E. C. (eds) Dinosaurs Past and Present, Volume II. Natural History Museum of Los Angeles County/University of Washington Press (Seattle and Washington), pp. 82-99.
Harris, J. M. 1987. Introduction. In Czerkas, S. J. & Olson, E. C. (eds) Dinosaurs Past and Present, Volume I. Natural History Museum of Los Angeles County/University of Washington Press (Seattle and Washington), pp. 1-6.
Lauber, P. 1989. The News About Dinosaurs. Bradbury Press, New York.
Wallace, J. 1989. The Rise and Fall of the Dinosaur. David & Charles, Newton Abbot, London.
The Life Appearance of Sauropod Dinosaurs
If you’re a regular TetZoo reader you’ll be familiar with my several articles on the life appearance of Mesozoic dinosaurs, an issue in which I have a special interest. Alas, several of these articles were published at TetZoo ver 2 – the ScienceBlogs years – and hosting issues at the site concerned mean that they’re currently appearing sans all of the many images I so lovingly uploaded. Which is a pain.

A really nice, life-sized model of a titanosaurian sauropod, on display in Romania and constructed by Brian Cooley. Image: Darren Naish.
Partly as a consequence, and partly because the issue is on my mind due to several concurrent projects (he says, cryptically), now is a good time to talk once more about the life appearance of Mesozoic dinosaurs, and I’m going to start with sauropods. Sauropods have been covered quite a few times on TetZoo before, as you can see from the list of links at the bottom of this article.
The head, and face especially. We’ll start with the head. Sauropod skulls were proportionally small compared to the overall size of the animals, but not comically so. It’s also worth saying that their eyes – while pretty big in absolute size (based on the size of the eye socket and the sclerotic rings preserved in fossils) – are easy to over-emphasise in illustrations. If you’ve heard that sauropods might have had trunks and thought it reasonable or plausible… well, it’s a poor idea that’s neither reasonable nor plausible, and has a lot counting against it, as covered in the ver 3 article here.

Did sauropods have trunks? NO. It’s the dumbest idea ever, and every piece of evidence counts against it. Several authors have independently explored this idea, including Robert Bakker (upper right) and Bill Munns (lower right). The two images at left are from Knoll et al.’s (2006) paper refuting the trunk idea. Images: Knoll et al. (2006), Bakker (1986), Bill Munns (original here).
Trunks are a no, but did sauropods have ‘lips’ or ‘cheeks’? A whole article could be written on this issue. For now, I’ll summarise things by saying that sauropod skull bone texture indicates that they likely did have extra-oral tissues that mostly covered their teeth (Witton 2018), meaning that they were facially similar to lizards and kin.
A suggestion that some titanosaurs had blade-like cutting sections in the posterior sections of their jaws – so-called ‘guillotine crests’ (Apesteguía 2004) – looks unlikely given that it would require the animals concerned to do their food procurement at the sides of the jaws, rather than at the front like all other sauropods and other dinosaurs. It’s more likely that a mistake has been made here and that the jaw segments thought to support those blade-like, keratinised sections were, while sharp-edged, covered in normal lip tissue. The possibility that diplodocoids and maybe other sauropods too might have had true beaks at the font of the mouth has recently been put forward, but so far only in preliminary fashion.

Apesteguía (2004) argued that some titanosaurs - this is the rhino-sized Bonitasaura - had keratinised ‘guillotine crests’ on the edges of the jaws. I think that this is likely a mistake. Image: Apesteguía (2004).
The nostrils and nose. As is now widely known, and near-universally accepted, the external or fleshy nostrils of sauropods were almost certainly not located way up on the forehead, far from the front of the snout, as was long thought based on the retracted position of the bony nostril openings. A more ‘normal’, anterior position for the nostrils was demonstrated by Witmer (2001) who pointed to evidence from blood vessel and nerve impressions and associated cranial hollows, all of which are located on the anterior part of the snout. These indicate the most likely position of the fleshy nostril and associated blood vessel clusters.

The soft dinosaur revolution comes to Sauropod Town. At left, an image by Matt Wedel which shows why we need more soft tissue on our sauropods. At right, Larry Witmer’s (2001) depiction of the different possibilities as goes nostril position in sauropods. (a) is the most likely option based on anatomical data. Image: Mathew Wedel, Witmer (2001).
There are also reasons for thinking that the giant, cavernous bony nostril openings and tall nasal bars of some macronarian sauropods supported, and were surrounded by, dome-shaped soft tissue convexities, superficially recalling the bulging nasal regions of some living monitor lizards. Part of my reason for saying this comes from the nasal anatomy of the exquisitely preserved South American titanosaur Sarmientosaurus. Here, an anteriorly projecting bony spine located along the midline and associated bar-like structures on the lateral edges of the large narial fossa – the big bony opening surrounding the bony nostril – indicate that a wide, convex mass of tissue connected the forehead with the sides and front of the snout’s upper surface (Martínez et al. 2016). Sarmientosaurus isn’t unique in this respect but is a particularly good example.

The skulls of some sauropods - this is the titanosaur Sarmientosaurus - indicate that there were bulbous nasal structures covering much of the snout region. Image: WitmerLab.
Indeed, skull openings in general were almost definitely not sunken in appearance or otherwise all that obvious, something that’s being said of archosaurian faces in general as artists and anatomists have learnt to take better attention of the conditions present in living animals (where cranial openings are just about never obvious in the live creature).
Accordingly, sauropod faces were seemingly ‘softer’ and more ‘padded’ than convention would have it, at least some of – perhaps all of – the nasal and forehead region being convex and fleshy, rather than shrink-wrapped and covered in thin skin alone (Witton 2018).
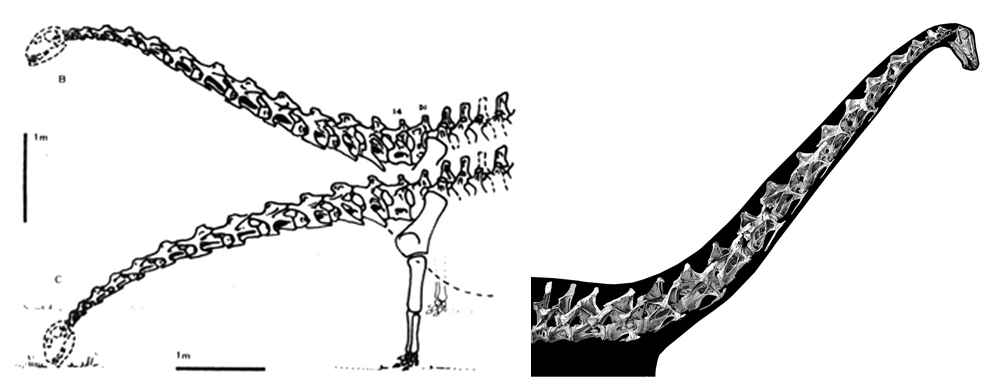
Were sauropod necks mostly semi-horizontal and with a limited range of motion, as argued by Martin (1987) and illustrated at left? Nope; it’s more likely that the necks were often held aloft and far more flexible, as argued by Taylor et al. (2009) and depicted at right. Images: Martin (1987), Taylor et al. (2009).
The neck. The most remarkable feature of sauropod anatomy is the neck. As goes how the neck was held and how flexible it was, several possibilities have been put forward, some workers arguing that it was held in a mostly horizontal attitude for much of the time and with only a limited range of lateral and vertical flexibility. I’m part of a group who argue for mostly elevated habitual neck poses (even in diplodocids) and a wide range of flexibility (Taylor et al. 2009). Arguments that sauropods must have been horizontal-necked do not take account of the flexibility permitted by cartilage, or – in living animals of all sorts – the ranges of motion that happen at zygapophyseal junctions and at the neck base and head-neck junctions.

There are good reasons for thinking that sauropod necks were habitually inclined upwards, as is typical for terrestrial tetrapods in general, and several fossils are actually preserved with the neck in this orientation. This montage by Greg Paul shows several of the fossil concerned. Image: Paul (1998).
It's been convention to show the sauropod neck as a featureless tubular structure, like a hose. This is mostly wrong, in part because the shapes of the vertebrae reveal a more unusual cross-sectional shape that varies from one sauropod group to the next. In some sauropods (like mamenchisaurs), the vertebrae are quite narrow and the neck would have looked laterally compressed in places, as it is (for at least some of its length) in giraffes. In diplodocoids – apatosaurines in particular – the neck is narrow close to the head but wide for much of its length, and subtriangular in cross-section, being widest across the neck’s underside. A neck that was quite narrow in its anteriormost quarter or so but was markedly wide for the rest of its length seems to have been the case in brachiosaurs and at least some titanosaurs.

The necks of some sauropods - a good example is the ultra-long-necked Omeisaurus, photographed at Zigong Dinosaur Museum - are somewhat laterally compressed for at least part of their length. Image: Darren Naish.
Indeed, the very base of the neck is remarkably broad in some sauropods – Camarasaurus is the classic example, where the neck base is not that different in width from the front of the chest – meaning that the neck would taper gradually along its length if you were looking at the animal from the front or back, or from above or below.

The neck bases of some sauropods - this is the Camarasaurus replica on display in London’s NHM - are shockingly broad. It would be wrong to show the neck as a narrow, hose-like object with parallel sides. Image: Darren Naish.
The vertebrae themselves are very complicated with large hollows on their sides, projecting neural spines on the apices and so on. In the most extreme version of the shrink-wrapping meme promoted by one or two palaeontologists and palaeoartists, sauropod necks have been depicted as if these structures should be visible in the live animal, Ely Kish’s apatosaurines from 1983 being the ultimate example. This was definitely not the case. It remains uncertain how much musculature and soft tissue surrounded the vertebrae, but it was almost certainly (based on the anatomy of living animals) enough to obscure the form of the vertebrae, their bulbous junctions perhaps being visible in sauropods of some or many sorts.

The brilliant, late Ely Kish, one of the best palaeoartists of all time, constructing a scale model of Apatosaurus (in preparation for a large painting). Partly on the advice of palaeontologist Dale Russell, she depicted great lateral concavities on the sides of the neck. Image: Russell (1987).
Necks as display banners. The unprecedented form of the sauropod neck makes it plausible – perhaps even likely – that the neck was used as a display structure. Phil Senter’s suggestion that the size and length of the neck was driven, in evolutionary terms, by its use as a display structure (Senter 2007) is not supported by evidence (Taylor et al. 2011). However, this doesn’t discount co-option of the neck in display, so it really is – while wholly speculative right now – worth taking seriously the possibility that display structures of various kinds could well have adorned sauropod necks. These could include wattles, dewlaps, spiky frills, spines, filaments, inflatable pouches or distensible flags or flaps. A few artists have explored these possibilities, most notably Brian Engh and Emiliano Troco.

As weird as it might seem, it is at least plausible that the sauropod neck was decorated with weird soft-tissue display structures, as depicted here on a diplodocid. Image: Emiliano Troco.
Hands. Sauropod hands are extremely odd, and very different from the rounded, elephant-style structures, edged with big nails and/or several claws, shown so often in artistic reconstructions and museum models. Indeed, they’re sufficiently weird and interesting that I’ve written whole articles about them before but, as I said above, these are currently lacking all of their relevant illustrations and are thus all but useless.

Sauropod hands are essentially unique. They’re weird, semi-tubular structures with pillar-like metacarpals. At left, a brachiosaur hand. At right, the hand of the turiasaur Zby. Images: Anthony Maltese, Darren Naish.
The sauropod hand is essentially a semi-tubular structure formed of elongate metacarpals arranged, pillar-like, in a semi-circle. The posterior surface – corresponding to the palm – was hollow, the consequence being a semilunate area of contact with the ground. We’ve known since at least 1940 that sauropod hands had this very unusual form thanks to fossil tracks (Falkingham et al. 2014), which makes it all the weirder that people have ignored this information and merrily continued giving sauropods elephant-like hands across the decades.

Roland T. Bird’s sauropod track illustrations from the 1940s - shown here - clearly show the true, highly unusual form of the sauropod manus. Shame on those who ignored this information in the following decades. Image: Falkingham et al. (2014), CC BY 4.0.
Tracks also show that nails and claws were absent from the hands, except on the thumb where a pointed claw – which varied considerably in exact shape and size from one group to the next – projected inwards and slightly backwards. The thumb claw was lost within Titanosauria, meaning that at least some members of this group lacked nails and claws on their hands altogether. I will concede that poorly defined convexities corresponding to manual digits were present in at least some sauropods, but they still wouldn’t have looked like distinct digits.
Some tracks appear to show that tough semi-conical tubercles projected from the skin on the front and sides of the hand, perhaps giving part of the hand a spiky or tuberculated appearance (Milàn et al. 2005). Maybe these structures had a role in foraging, digging, display or combat, since it’s plausible that they made the hands gnarlier and harder than they would have been otherwise.

Vertical scores associated with sauropod hand prints indicate that at least some of them had tuberculate hand skin, as depicted here (at right) in this modified version of a Greg Paul illustration. Image: Milàn et al. (2005).
Feet. Three large, curved claws projected anterolaterally from the inner three toes. At least some sauropods possessed four such claws. The outer two toes projected as blunt, rounded convexities in some, most or all sauropods. This is obvious from at least some Brontopodus tracks (Meyer et al. 1994). They might have had nails but were more likely lacking horny structures of any sort and essentially continuous with the rest of the foot’s outer surface. In contrast to the hand, the foot did have a massive fatty pad at its back.
Until recently it was thought that the metatarsals were held at a high angle, the consequence being a short foot shaped like that of an elephant but for the claws (Paul 1987). Data from articulated skeletons and tracks, however, show that the metatarsus was not as erect as argued, in which case the toes were longer and flatter than depicted by some artists.

Tschopp et al. (2015) used data from complete Camarasaurus hands and feet to produce the skeletal and soft-tissue models you see here (in 1 and 2), and then used these to generate tracks (3). The tracks are an exact match for real fossil tracks. Image: (c) Tschopp et al. (2015).
On the subject on feet and limbs, trackways also show that sauropods differed in how they placed their hands and feet, some walking with a very narrow gait (the hands and feet being placed close to the midline), others using a wide gait (where there was some short distance between the hands and feet of the left side versus those of the right), and others being intermediate. It should be remembered that even the widest-gauge sauropods did not walk with their feet all that far apart, but that the legs were almost certainly angled inwards, as is typical in living animals. For more on this issue see Scott Hartman’s article here.
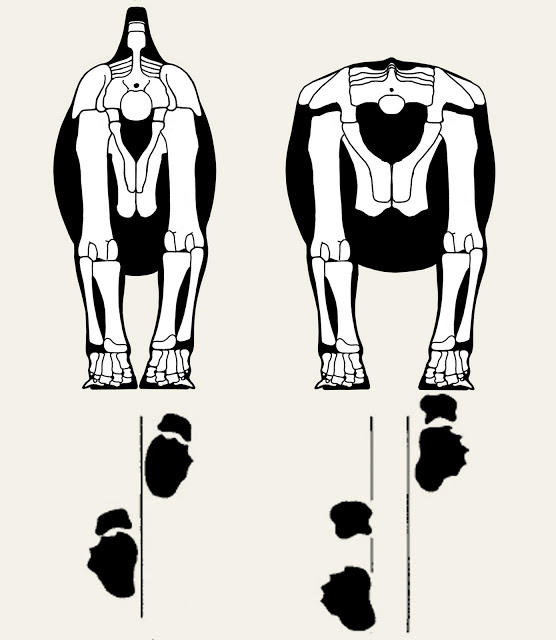
Sauropods were variable in cross-sectional shape and in whether they walked with narrow-gauge or wide-gauge gaits, as depicted here by Scott Hartman. Image: Scott Hartman’s skeletaldrawing.com
The body and tail. Sauropod bodies were variable in cross-sectional shape, length and other details. Diplodocoids, for example, were relatively narrow, deep-bodied and with a tall ridge formed from their neural spines running along the back, while titanosaurs were extremely broad across the hips and must have been just about flat across the back. The thorax may have sloped down ever so slightly in diplodocoids (in part because their forelimbs were shorter than their hindlimbs) whereas the thorax was angled upwards slightly or even markedly in some macronarians, like brachiosaurs and some titanosaurs.
Articulated skeletons show that the tail mostly projected horizontally from the pelvis (a subtle arch at the tail base is present in diplodocoids and some others, in fact), but the macronarians with the upward-sloping bodies also had a downward-sloping tail. A real curiosity is provided by the tails of some mamenchisaurs which seem to have projected upwards at an angle. This has been discussed and illustrated by Paul (2010) and more recently by Hallett & Wedel (2016), but has otherwise gone undiscussed as far as I know. It sounds so odd that surely some mistake has been made… though I really don’t think it has.

Tall neural spines, massive, wing-shaped transverse processes and other structures show that an enormous quantity of musculature would have been obvious along the proximal part of the sauropod tail. This is the tail of … ugh .. Dippy, the cast of Diplodocus carnegii until recently on show in London. Image: Darren Naish.
As is typical for non-bird dinosaurs, the musculature at the base of the tail was (so we can say from the relevant bony attachment points) evidently enormous and bulky, the proximal part of the tail likely being similar in width to the pelvis and thighs combined.
The integument. We know for definite that sauropods of all groups were scaly animals, since scaly skin impressions and actual preserved skin patches are known for diplodocids, Camarasaurus and titanosaurs at least. A few skin folds here and there were almost certainly present (say, where the limbs met the body, at the limb joints, and at mobile zones in the neck) but a fissured, wrinkled or scored elephant-like skin is a big fat no and every effort should be made to avoid it in artistic depictions.

Haestasaurus from the English Wealden (shown at left) and a few other sauropods preserve polygonal scales that vary somewhat in size. Image: Darren Naish, Czerkas (1994).
Polygonal – specifically, pentagonal, hexagonal and heptagonal – scales are known for the macronarian Haestasaurus and some diplodocids, while rounded scales possessing a papilliform texture of tiny bumps are also preserved in a diplodocid specimen (Czerkas 1994). The polygonal structures varied in size somewhat, those present in more mobile parts of the skin (say, the inner crease of the elbow) being smaller than those present in non-mobile sections. Even so, the largest scales were, at most, 60 mm across, which is not large at all on an animal more than 15 m long. This is pretty common in non-bird dinosaurs, by the way. The scales were often so small that they wouldn’t be readily visible from any distance greater than a few metres. The papilliform texture on those diplodocid scales seems to have been widespread across sauropods and means that sauropod skin would have been rough to the touch.
A diplodocid specimen from Wyoming that has scaly skin preserved also preserves tall, conical dermal spines (note: they are not horn-covered bony structures) that must have been arranged along the dorsal midline (Czerkas 1992). These are variable in height (the biggest are 18 cm tall) and seem to have formed a single row along the top of the tail’s proximal part at least. This specimen was originally implied to belong to Diplodocus but is of indeterminate identity. It might belong to Kaatedocus, since remains of this dinosaur come from the exact same quarry.

Large, laterally compressed conical and semi-conical dermal structures lined the upper surface of the tail (at least) in some diplodocids, and perhaps in other diplodocoids and sauropods too. Image: Czerkas (1994).

The conical and semi-conical dermal spines of some diplodocids were variable in height, breadth and shape, as illustrated here. Image: Czerkas (1994).
Were these structures present across all diplodocids, all diplodocoids, all neosauropods or even all sauropods, or were they exclusive to one small clade, perhaps even just to Kaatedocus itself? In the absence of further information we can’t say, but it’s appropriate to depict them on diplodocids and their close kin at least, in the absence of further information. The possibility that shorter and/or taller dermal structures of this sort were present elsewhere on other sauropods exists.

The discovery of those conical and semi-conical spines in a diplodocid led Stephen Czerkas to produce this new look for these dinosaurs. It might be correct, but it is not clear whether the structures were as extensive along the animal’s length as shown here. Image: Czerkas (1992).
Horn-covered osteoderms - that is, lumps and nodules with a bony core - were arranged across the backs and flanks of some titanosaurs, specifically the members of the clade Lithostrotia. These structures were variously rounded or oval, sometimes quite flat and sometimes slightly or strongly domed. They appear to have been numerous and prominent in some of the relevant species and would have given their backs and sides a pebbly, armoured appearance.
Colours. There’s a traditional view stating that big dinosaurs were likely grey and plain because big living mammals are, as is the Komodo dragon, I suppose. I’m going to be bold here and say that there’s no reason whatsoever to take any notice of this. Sauropods were not mammals or Komodo dragons. While an argument could be made that camouflage would have been beneficial, and that certain pigments and patterns would have been helpful or necessary for physiological reasons (heat-shedding, heat retention, UV protection and so on), bold and complex patterns and bright, even vibrant colours are all consistent with the ecophysiological demands of being a sauropod, and the good colour vision and complexity of sauropod integument, and likely reliance on visual display, mean that they very likely could have been more like gigantic lizards or birds than elephants. In any case, giraffes, perenties and others show that big, terrestrial animals don’t have to be bland and grey as has often been stated.

The idea that big terrestrial animals have to be bland never was correct. Perenties Varanus giganteus might not be that colourful, but they help emphasise the possibility that striking patterns can be present even in very large terrestrial reptiles. Image: (c) Stephen Zozaya.
In the absence of any direct evidence, it seems reasonable to me to reconstruct barred, striped, reticulated or dappled colour schemes, to depict bright colours on faces, necks or other areas considered relevant to display, and to imagine any colours reasonable for big, terrestrial animals associated with woodlands, parks, scrubby places, mangroves and all the other habitats frequented by this long-lived, diverse group of animals.

Want to know more about sauropod life appearance? It sometimes surprises people that there are scarcely any good books dedicated to specific Mesozoic dinosaur groups. For sauropods, the main must-have volume is Mark Hallett and Matt Wedel’s The Sauropod Dinosaurs. It’s really good. Dinosaurs Past and Present Vol II is also worth getting, in part because it includes Greg Paul’s (now quite dated, but still useful) article on dinosaur life appearance. Images: amazon (here and here).
If we really want to imagine sauropods as living animals, there is – of course – so much more to say. Body language, posture, gait, social life, anti-predator responses, feeding behaviours, digestive function, sleep and so much more are all things that need to be considered. But that’s where I’ll stop for now. I hope you found this interesting, and we’ll be looking at dinosaur life appearance again sometime soon.
For previous TetZoo articles on sauropods, see…
The hands of sauropods: horseshoes, spiky columns, stumps and banana shapes, October 2008
Sauropod dinosaurs held their necks in high, raised postures, May 2009
Necks for sex? No thank you, we're sauropod dinosaurs, May 2011
The sauropod viviparity meme, May 2011
The Second International Workshop on the Biology of Sauropod Dinosaurs (part I), December 2011
The Second International Workshop on the Biology of Sauropod Dinosaurs (part II), January 2012
Junk in the trunk: why sauropod dinosaurs did not possess trunks, November 2012
The changing life appearance of dinosaurs, September 2014
Brian J. Ford's Aquatic Dinosaurs, 2014 Edition, February 2015
That Brontosaurus Thing, April 2015
10 Long, Happy Years of Xenoposeidon, November 2017
Refs - -
Apesteguía, S. 2004. Bonitasaura salgadoi gen. et sp. nov.: a beaked sauropod from the Late Cretaceous of Patagonia. Naturwissenschaften 91, 493-497.
Czerkas, S. A. 1992. Discovery of dermal spines reveals a new look for sauropod dinosaurs. Geology 20, 1068-1070.
Czerkas, S. A. 1994. The history and interpretation of sauropod skin impressions. Gaia 10, 173-182.
Knoll, F., Galton, P. M. & López-Antoñanzas, R. 2006. Paleoneurological evidence against a proboscis in the sauropod dinosaur Diplodocus. Geobios 39, 215-221.
Martin, J. 1987. Mobility and feeding of Cetiosaurus (saurischia, sauropoda [sic]) - why the long neck? In Currie, P. J. & Koster, E. H.(eds) Fourth Symposium on Mesozoic Terrestrial Ecosystems, Short Papers. Boxtree Books (Drumheller, Alberta), pp. 154-159.
Meyer, C. A., Lockley, M. G., Robinson, J. W. & dos Santos, V. F. 1994. A comparison of well-preserved sauropod tracks from the Late Jurassic of Portugal and the western United States: evidence and implications. Gaia 10, 57-64.
Milàn, J., Christiansen, P. & Mateus, O. 2005. A three-dimensionally preserved sauropod manus impression from the Upper Jurassic of Portugal: implications for sauropod manus shape and locomotor mechanics. Kaupia 14, 47-52.
Paul, G. S. 1987. The science and art of restoring the life appearance of dinosaurs and their relatives - a rigorous how-to guide. In Czerkas, S. J. & Olson, E. C. (eds) Dinosaurs Past and Present Vol. II. Natural History Museum of Los Angeles County/University of Washington Press (Seattle and London), pp. 4-49.
Paul, G.S. 1998. Terramegathermy and Cope’s rule in the land of titans. Modern Geology 23, 179-217.
Paul, G. S. 2010. Dinosaurs: A Field Guide. A & C Black, London.
Russell, D. A. 1987. Models and paintings of North American dinosaurs. In Czerkas, S. J. & Olson, E. C. (eds) Dinosaurs Past and Present, Volume I. Natural History Museum of Los Angeles County/University of Washington Press (Seattle and Washington), pp. 114-131.
Senter, P. 2007. Necks for sex: sexual selection as an explanation for sauropod dinosaur neck elongation. Journal of Zoology 271, 45-53.
Taylor, M. P., Hone, D. W. E., Wedel, M. J., & Naish, D. 2011. The long necks of sauropods did not evolve primarily through sexual selection. Journal of Zoology 285, 150-161.
Taylor, M. P., Wedel, M. J. & Naish, D. 2009. Head and neck posture in sauropod dinosaurs inferred from extant animals. Acta Palaeontologica Polonica 54, 213-220.
Witmer, L. M. 2001. Nostril position in dinosaurs and other vertebrates and its significance for nasal function. Science 293, 850-853.
Heilmann, Thompson, Beebe, Tetrapteryx and the Proavian
Many people familiar with ideas on the early evolution of birds and of bird flight will know of the Proavis or proavian, a hypothetical bird ancestor illustrated and discussed by William Pycraft (1868-1942) in 1906 but made better known by Gerhard Heilmann (1859/1861-1946) during the 1910s and 20s. Versions of this creature were later illustrated in colour by Zdeněk Burian for popular books of the 1960s, 70s and 80s. In the article that you’re about to read, we’re predominantly interested in Heilmann’s take on the Proavis.

Caption: fighting, gliding and climbing proavians, depicted in a cluttered woodland environment, by Gerhard Heilmann. This is typical of Heilmann’s many excellent scenes. He used posed plaster models of the creatures before drawing. This illustration was used in the 1916 Danish version of the book (it first appeared in one of his 1912 articles) but does not appear in the 1926 English version. As we’ll see below, the decision to exclude some images from the 1926 version might have served to make it seem more scholarly than its predecessor. Image: Heilmann (1916).
Heilmann was an artist and graphic designer by profession but his interest in bird evolution and anatomy was such that he published a series of articles on the subject for the Danish Ornithological Society between 1912 and 1916 (Nieuwland 2004, Ries 2007).

Caption: self-portrait of Gerhard Heilmann, produced in 1902. Heilmann was an argumentative man often in conflict with “various forms of authority”, and at loggerheads with his own family (Nieuwland 2004). This illustration was included in Nieuwland (2004).
Compiled, these articles described Heilmann’s take on the diversity of ancient fossil birds, embryological development in birds and reptiles, and bird anatomy, with the last in the series explaining what these data meant for the appearance and lifestyle of his Proavis, an imagined animal older and anatomically more archaic than Archaeopteryx. These articles were published together in a Danish-language book Vor Nuvaerende Viden om Fuglenes Afstamning (Heilmann 1916), the title meaning Our Current Knowledge of the Descent of Birds.
Heilmann’s The Origin of Birds. This body of work was sufficiently impressive that – while Heilmann was mostly shunned in Denmark – it was well received elsewhere, and he was encouraged by scientists in Germany in particular to translate it into English. The result was the 1926 The Origin of Birds (republished in the USA in 1927), a scholarly and well-illustrated book that was enthusiastically accepted internationally and almost immediately deemed the most authoritative work on bird origins. The 1926 volume is quite different from that of 1916 in the illustrations it includes (as we’ll see below), but also in how scientific and speculative the text is, some more fanciful sections on palaeobiology being absent from the English version.
It has been said several times that Heilmann’s work was deemed so impressive that, rather than inspire new work on the subject, it effectively bought research on bird origins to a halt. Most relevant workers (albeit not all) now considered the question of bird origins to be resolved. As noted by historian Ilja Nieuwland, things might not have gone this way had the English-speaking readers of Heilmann’s book known that he was an amateur scientist and an artist by trade (Palm 1997, Nieuwland 2004).

Caption: Heilmann’s draftmanship was superb. This illustration (fig. 140 from Heilmann 1926) shows thigh feathering on the chicks of various birds, with 8 being a gliding gecko. “The animal with which we may best compare the bird-ancestor, is the Fringed gecko” [sic] (p. 197). Image: Heilmann (1926).
Heilmann thought that bird ancestors were likely quadrupedal gliders, similar superficially to living gliding geckos (Heilmann 1926, p. 197). This statement is somewhat odd in view of his endorsement elsewhere of a more cursorial view of these animals, but it seems that he imagined them adopting very different poses when on the ground versus when climbing. Heilmann also argued that birds were not dinosaurs – despite his good understanding of the compelling anatomical similarity between theropod dinosaurs and birds – but were instead the descendants of a group closely related to – and supposedly ancestral to – dinosaurs, termed ‘pseudosuchians’*. This was because of his adherence to ‘Dollo’s Law’ – the (erroneous) idea that a lost anatomical structure cannot be regained – and his mistaken contention that dinosaurs lacked clavicles. If dinosaurs couldn’t be ancestral to birds, the true ancestors must have been another group, hence Heilmann’s use of the more archaic ‘pseudosuchians’, already mooted as possible bird ancestors by Robert Broom in his 1913 description of the South African Euparkeria.
* The term pseudosuchian is today applied to the archosaur lineage that includes crocodylians and all of their extinct relatives. Given the historical baggage that comes with the term, I personally don’t think that this is a good idea at all and would prefer it if another name were used for the lineage concerned (like Crurotarsi)… but that’s an issue for another time.

Caption: Euparkeria capensis has long been intimated as a sort of bird ancestor by those looking for such creatures outside of theropod dinosaurs. Not only is this animal only very distantly related to birds, it’s not even part of the crown-archosaur clade. Image: Taenadoman, CC BY-SA 3.0 (original here).
By combining features common to Archaeopteryx as well as to the ‘pseudosuchians’ Aetosaurus, Euparkeria, Ornithosuchus and Saltoposuchus, Heilmann (1916, 1926) described how he invented a creature that looked somewhat like a theropod but had a more archaic skull and foot, retained a fourth metacarpal, and had a smaller pelvis with far shorter pubic and ischial bones.

Caption: Heilmann’s skeletal reconstruction of Proavis, as depicted in the 1916 Danish version of the book… but not in the 1926 English version. Note the lack of long feathers on the hindlimbs. Incidentally, note also that Heilmann was partly responsible for encouraging the belief that forelimb feathers did not cover the hands during the earliest stages of bird evolution. Image: Heilmann (1916).
Heilmann, Thompson and D’Arcy Thompson grids. It’s a matter of some interest that Heilmann didn’t just guess what the imagined morphology of the proavian would be, as might be assumed given the English edition of his book. Instead, he used a technique whereby the relevant anatomical regions were mapped on to a grid and then distorted to mimic the evolutionary process. This grid-based deformation process was pioneered by D’Arcy Wentworth Thompson (1860-1948) and was explained most thoroughly in his 1917 book On Growth and Form (Thompson 1917). It was actually put forward beforehand in 1915 (Thompson 1915).

Caption: Thompson applied his Cartesian grid technique to many animal lineages, his aim being to show that mathematically predictable transformation in one or more anatomical regions could result in profound anatomical change. Here’s one of my favourite examples: how you can derive a molid sunfish (Mola was known to Thompson as Orthagoriscus) from a porcupinefish. Image: Thompson (1917).
By placing grids on top of diagrams of related animals (say, the skull of a human and a chimp), Thompson showed how the skewing of the grid in a certain direction “would result in the sort of changes that would allow the emergence of a new species” (Naish 2017, p. 116). These grids are generally termed D’Arcy Thompson transformation grids, Cartesian transformations or Cartesian grids, and their use was quite popular in the evolutionary literature of the early 20th century. Heilmann was inspired to use the technique after Thompson wrote to him about bird evolution in 1915, and the two wrote to each other on many occasions about the technique and its application to ideas on the evolution of horses, hominids and birds. They also discussed how the resulting illustrations could be used in Thompson’s On Growth and Form (Ries 2007).

Caption: Heilmann included these Cartesian transformations in the 1916 version of his book, but they weren’t included in the 1926 English version. His take on Proavis was not, therefore, simple guesswork. Image: this montage is from Witmer (1991) but the originals are from Heilmann (1916).
Heilmann’s commitment to the technique is demonstrated by the fact that the 1916 Danish edition of his book includes grids that depict the inferred evolution of the avian skull, forelimb and pelvis (I don’t own a copy of the Danish edition, but the diagrams are included in both Witmer (1991) and Ries (2007)). With Ornithosuchus and Euparkeria as ‘starting points’ and Archaeopteryx and modern birds as ‘end points’*, Heilmann used the grids to create relevant intermediates, the results allowing him to generate, piecemeal, his Proavis (Heilmann 1916). A lateral view of the hypothetical animal – showing it in a neural, non-dynamic walking pose – was included in the Danish edition (Heilmann 1916, Ries 2007).
* It should be noted that Heilmann’s Archaeopteryx was not accurate, some of its details (in the skull especially) being semi-hypothetical and informed by Heilmann’s interpretation of ‘pseudosuchians’ like Aetosaurus.

Caption: the more dynamic, climbing version of Heilmann’s Proavis skeletal reconstruction, the only version of the reconstruction included in the 1926 English version of his book. Image: Heilmann (1926).
Remarkably, none of this was included in the English edition at all, this creating the impression that Heilmann simply invented the proavian via intuition and guesswork. He didn’t: it was generated via a testable, repeatable method (albeit using flawed anatomical data). Furthermore, the fact that Heilmann’s skeletal reconstruction of Proavis included in the English edition (Heilmann 1926) is the more dynamic climbing version of the creature gives it a less scientific, more speculative, artistic air than the walking version of 1916.
Heilmann, Beebe and Tetrapteryx. When it came to feathering and other aspects of the integument, Heilmann noted his thoughts on another Proavis-type animal, this time the one invented by Charles William Beebe in 1915. Beebe (1877-1962) was an interesting person. He was a naturalist, ecologist, ornithologist, marine biologist, author and explorer, among other things. Today, he might be best known for the deep-water observations he made while in a bathysphere off the coast of Nonsuch Island off Bermuda during the 1930s.
![William Beebe’s hypothetical tetrapteryx creature. A pre-Archaepteryx glider. You’ll note that Beebe was a pretty good artist [UPDATE: this is an error. The illustrations in this publication were by Dwight Franklin, not by Beebe!]. Image: Beebe (191…](https://images.squarespace-cdn.com/content/v1/510be2c1e4b0b9ef3923f158/1546356482204-OC8JSKSSMTH2YL41VQNL/Heilmann-Proavian-Beebe-Tetrapteryx-Jan-2019-Tetrapod-Zoology.jpg)
Caption: William Beebe’s hypothetical tetrapteryx creature. A pre-Archaeopteryx glider. You’ll note that Beebe was a pretty good artist [UPDATE: this is an error. The illustrations in this publication were by Dwight Franklin, not by Beebe!]. Image: Beebe (1915).
When it came to bird origins, Beebe thought that birds originated via a ‘tetrapteryx’ phase where large feathers on the proximal hindlimb were present and functioned in slowing descent during leaping and gliding (Beebe 1915). Both the fossils of Archaeopteryx and the nestlings of living bird species demonstrated the antiquity of these ‘pelvic wings’, Beebe (1915) said. Heilmann (1926) disagreed. His own examination of bird nestlings pertaining to bird lineages across the family tree resulted in his “complete disappointment, for what I found was wholly negative; there was not in any of them the slightest trace of a “pelvic wing”” (p. 194).
As for Archaeopteryx, Heilmann (1926) argued that the hindlimbs were arranged such that they could never have had an aerodynamic function, nor was there evidence for long hindlimb feathering of the sort consistent with ‘pelvic wings’. Ergo, Heilmann illustrated Archaeopteryx without long hindlimb feathering, and he didn’t think that Proavis had them either: his skeletal reconstructions (both the 1916 walking version and 1926 climbing one) lack them (the 1926 climber has short hindlimb feathers, similar in length to those elsewhere on the body) and the gliding proavians included in his various drawings clearly lack them as well. His most famous proavian figure – the semi-erect one standing in a conifer tree (shown below) – does have long feathers on the rear edge of the thigh, though they certainly aren’t as long as they should be as per Beebe’s tetrapteryx model.
The iconic tree-dwelling proavian. The iconic tree-dwelling proavian I have in mind here is one of Heilmann’s best-known and most frequently reproduced illustrations. It has what appears to be great symbolic significance if you’ve read the English version of The Origin of Birds since it’s the very last image in the entire book (Heilmann 1926, p. 199), thus appearing as a visual metaphor for an imagined phrase: “Finally, I can reconstruct the real ancestor of birds, and it looked like this”.

Caption: Heilmann’s most iconic version of his Proavis. Note the long feathers on the back of the thigh and how they’re invisible and thus not deemed important in the gliding individual in the background. Heilmann said that the long feathers close to the base of the tail would produce the required lift in this region. Image: Heilmann (1926).
I was surprised to learn that Beebe produced what looks like his own version of this illustration, and inserted it as a plate in a book published in 1915. However, the image itself is not dated 1915 and does not definitely pre-date Heilmann’s illustration. My suspicion – and that of Paul Stewart, who kindly brought my attention to the image – is that Beebe copied Heilmann’s proavian for his own personal use (the illustration was not intended for publication*). This in itself is interesting since it could mean that Beebe personally endorsed Heilmann’s view of Proavis and regarded it as a ‘descendant’ of his tetrapteryx creature… which it wasn’t, given that Heilmann rejected Beebe’s idea, as we saw earlier. Again, my thanks to Paul Stewart for sharing this image with me and permitting its use here.
* Thanks to Jonathan Kane, I’ve learnt that it was previously published in Tim Berra’s 1977 William Beebe: An Annotated Bibliography.

Caption: William Beebe seems to have produced his own take on Heilmann’s iconic proavian scene. We’re presuming that it was produced some time after Heilmann’s illustration was, but this hasn’t yet been fully confirmed. The illustration appears here courtesy of Paul Stewart.
Today, the significance of Beebe’s tetrapteryx idea is a bit uncertain and, frankly, depends on who you ask. Archaeopteryx probably did have long feathers on the hindlimbs (Longrich 2006), and they perhaps had an aerodynamic role if this animal indulged in aerial locomotion (which it likely did). Furthermore, the discovery of prominent ‘hindlimb wings’ in other feathered dinosaurs (most notably the Chinese dromaeosaur Microraptor) has led some experts to note the predictive power of Beebe’s suggestion (Kane et al. 2016) and to favour the presence and importance of hindlimb feathers in the earliest birds (Longrich 2006, Chatterjee & Templin 2007, Zheng et al. 2013, Xu et al. 2014). However, it’s not at all certain that large, aerodynamically ‘functional’ hindlimb feathers were present consistently across the lineages concerned or that they had the role that Beebe imagined (O’Connor & Chang 2015).

Caption: recent work shows that Archaeopteryx really did have long feathers on the proximal portions of its hindlimbs (these reconstructions are by Longrich (2006)), though they weren’t as long as the feathers Beebe imagined for his tetrapteryx stage creature. Image: Longrich (2006).
As always, there’s more to say and this article is already longer than intended. I also wanted to talk about Pycraft’s initial concept of the Proavis, and also Burian’s depiction, since he didn’t simply copy Heilmann’s illustrations but actually produced yet another hypothetical creature. These things will have to wait to another time. Also worthy of further discussion is the point made earlier about Heilmann’s influence happening despite his status as a ‘mere’ artist, a theme that has parallels elsewhere in the history of vertebrate palaeontology and is relevant to my writings on meme perpetuation, the scientific acceptance of feathering in non-bird dinosaurs and much else besides. I aim to explore these topics (and others) in future articles.
Caption: before Heilmann, there was Pycraft’s proavian of 1906. We’ll have to discuss this creature and its backstory another time. Image: Pycraft (1910).
For previous TetZoo articles relevant to the issues covered here, see…
Flight of the Microraptor, November 2013
The Integrated Maniraptoran, Part 1, May 2016
The Integrated Maniraptoran, Part 2: Meet the Maniraptorans, May 2016
The Integrated Maniraptoran, Part 3: Feathers Did Not Evolve in an Aerodynamic Context, May 2016
Naish and Barrett's Dinosaurs: How They Lived and Evolved, November 2016
Book Review: God's Word or Human Reason? An Inside Perspective on Creationism, March 2018
Refs - -
Beebe, C. W. 1915. A tetrapteryx phase in the ancestry of birds. Zoologica 2, 38-52.
Chatterjee, S. & Templin, R. J. 2007. Biplane wing planform and flight performance of the feathered dinosaur Microraptor gui. Proceedings of the National Academy of Sciences 104, 1576-1580.
Heilmann, G. 1916. Vor Nuvaerende Viden om Fuglenes Afstamning. Unknown publisher, Copenhagen.
Heilmann, G. 1926. The Origin of Birds. Witherby, London.
Longrich, N. 2006. Structure and function of hindlimb feathers in Archaeopteryx lithographica. Paleobiology 32, 417-431.
Naish, D. 2017. Evolution in Minutes. Quercus, London.
O’Connor, J. & Chang, H. 2015. Hindlimb feathers in paravians: primarily “wings” or ornaments? Biology Bulletin 42, 616-621.
Palm, S. 1997. The Origin of Flapping Flight in Birds. Svend Plam, Ballerop.
Pycraft, W. P. 1910. A History of Birds. Methuen & Co, London.
Ries, C. J. 2007. Creating the Proavis: bird origins in the art and science of Gerhard Heilmann 1913-1926. Archives of Natural History 34, 1-19.
Thompson, D. W. 1915. Morphology and mathematics. Transactions of the Royal Society of Edinburgh 50, 857-895.
Thompson, D. W. 1917. On Growth and Form. Cambridge University Press, Cambridge.
Witmer, L. M. 1991. Perspectives on avian origins. In Schultze, H.-P. & Trueb, L. (eds) Origins of the Higher Groups of Tetrapods: Controversy and Consensus. Cornel University Press (Ithaca, London), pp. 427-466.
Xu, X., Zhou, Z., Dudley, R., Mackem, S., Chuong, C.-M., Erickson, G. M. & Varricchio, D. J. 2014. An integrative approach to understanding bird origins. Science 346 (6215), 1253293.
Zheng, X., Zhou, Z., Wang, X., Zhang, F., Zhang, X., Wang, Y., Wei, G., Wang, S. & Xu, X. 2013. Hind wings in basal birds and the evolution of leg feathers. Science 339, 1309-1312.
Up Close and Personal With the Crystal Palace Dinosaurs
Like many of us, I have long enjoyed looking at the Crystal Palace dinosaurs and other prehistoric animal models, created in 1854, still on show more than 160 years later, and providing a remarkable showcase of ancient life as it was imagined at the time. But I’ve only ever seen them from afar. How fantastic would it be to examine them up close? Well…

Caption: you’ve seen the Crystal Palace dinosaurs before (or images of them, anyway), but you might not have seen them up-close like this. Neither had I prior to this very special visit. Image: Darren Naish.
Way back in September 2018, I was fortunate enough to attend the Crystal Palace Dinosaur Days event, part of the Heritage Open Days weekend occurring across the UK on the weekend concerned. I gave a talk and also led a tour around the prehistoric animal models (focusing on the reptiles and amphibians alone). Adrian Lister (of mammoth and Megaloceros fame) led a tour too, Mark Witton gave a talk on ‘Palaeoart After Crystal Palace’, and much else happened besides. I also have to mention the 3D-printed models of the dinosaurs made by Perri Wheeler. How I would love for these to be commercially available: I’m sure they’d be a success. So, it was a great event; well done Ellinor Michel and everyone else involved in the Friends of Crystal Palace Dinosaurs group (follow them on Twitter at @cpdinosaurs) for putting it together.

Caption: Perri Wheeler’s brilliant models of the three Crystal Palace dinosaurs (from back to front: Megalosaurus, Hylaeosaurus, Iguanodon). As a pathological collector of model dinosaurs, I sure would like to own a set, but I also sure would like for these models (or a set very similar to them) to be commercially available. Image: Darren Naish.
The real thrill, however, was not the talks nor the presence of the amazing and sometimes spectacularly good speakers but the fact that we were awarded special, up-close access to the prehistoric animal models. A dream come true. As you’ll know if you’ve visited Crystal Palace or read about it, the models are located on islands surrounded by a snaking waterway. In other words, they aren’t readily accessible. For the duration of Dinosaurs Days, however, a temporary bridge had been erected and – like Lord Roxton striding across a felled tree to Maple White Land – we made the crossing and stepped into a bygone era.

Caption: the amazing, enormous head of the Mosasaurus. Many of the scales on the body were recently repaired as the entire skin across the body was in a poor state. More on the mosasaur below. Image: Darren Naish.
Why erect a bridge to the islands in the first place? Both so that crucial landscaping and gardening can occur, and so that the models can be examined and evaluated for repair. They’re not in the best of shape, you see, and much work needs doing. Indeed, right now there’s a major push to get funding for a permanent bridge that will allow the continual access that’s required. This project only has a few days of fundraising left and there’s some way to go before the target is reached: go here and chip in if you can. You might have heard that the Mayor of London agreed to partly fund the project… as has legendary musician and song-writer Slash, since it turns out that he’s a big fan! I should add that Slash seems to be quite the fan of science in general, his twitter account revealing a definite tendency to use his powers for good.

Caption: the standing Iguanodon was given renovation and a new paint scheme within recent years. Unfortunately, further repair work is already required. Image: Darren Naish.
The reason I’m writing this article is not just to bring attention to this push for funding, but also to discuss and illustrate various of the remarkable details I got to see thanks to this up-close encounter. Before I start, be sure to read (if you haven’t already) the August 2016 TetZoo ver 3 article on the Crystal Palace models. Thanks to the Dinosaur Days event, I should add that I’ve been able to get hold of the guide that Richard Owen wrote to accompany the exhibition, or the 2013 reprinting (Owen 2013) of this 1864 publication (Owen 1854), anyway. It provides at least some background information on why the animals look the way they do.

Caption: head of the reclining Iguanodon. Only a privileged few have seen the head from its left side. Image: Darren Naish.

Caption: again, relatively few people will have seen the reclining Iguanodon from this side. It’s striking how natural, realistic and well-proportioned the model looks in this view: very much like a real animal. Image: Darren Naish.

Caption: Hylaeosaurus was thought by Owen and Hawkins to be an iguana-like reptile with a “lofty serrated or jagged crest, extended along the middle of the back”, though many aspects of the reconstruction were noted as being “at present conjectural” (Owen 2013, p. 18). Image: Darren Naish.
I’ll avoid repeating here the same points I made in my 2016 article but I will repeat my primary take-homes. Namely, that it’s disingenuous and naïve to criticise the models as outdated or as inaccurate, laughable follies, as is sometimes done. They have to be seen within the context of what was known at the time, there has to some acknowledgement of the fact that scientific knowledge has improved over time, and there should also be recognition of the fact that the models are more up-to-date than, and superior in technical accuracy and craftmanship to, the vast majority of modern efforts to portray prehistoric life. In the interests of correcting a mistake made in my 2016 article I should also point out that Crystal Palace is not in Sydenham as I stated, but in Penge. With that out of the way…

Caption: the pachydermal, vaguely bear-like Megalosaurus is actually a composite of information compiled from both Jurassic and Cretaceous theropods. The tall shoulder hump was included because Owen erroneously regarded the tall-spined Altispinax (previously Becklespinax) vertebrae as belonging to the shoulder region of Megalosaurus (Naish 2010). Image: Darren Naish.

Caption: a lot of detail was added to the megalosaur’s face, some of it is superficially crocodylian-like. Note the ominous cracks at the tip of the nose and along the side of the lower jaw. Image: Darren Naish.
It was a real thrill to see the remarkably detailed appearance of the three Crystal Palace dinosaurs: Iguanodon, Megalosaurus and Hylaeosaurus. Each has a very different skin texture, the Megalosaurus being the most unusual in that it doesn’t have the tile-like scales of the other two. Instead, it’s decorated with a crazy-paving-like covering. It’s not clear what Benjamin Waterhouse Hawkins (the designer and model-maker) was trying to achieve here since this is a rather non-reptilian look. Perhaps the aim was to give the animal a fissured skin texture vaguely like that of elephants.

Caption: view of the interior of the standing Iguanodon, with and without flash. The light at the far end of the image is coming in through the Iguanodon’s mouth. Image: Darren Naish.
Holes on the undersides of the Iguanodon and Hylaeosaurus mean that their insides can be inspected. As you can see from my photos, the models look like weird gabled roofs from the inside, numerous metal struts and poles helping to provide support. The hylaeosaur’s original head was removed since its weight was causing the model’s neck to break, and was replaced with a fibreglass copy. So, peer inside the hylaeosaur from beneath and you see the translucent interior of its face.

Caption: a view of the hylaeosaur’s interior! Image: Darren Naish.
Cracks, fissures and damaged sections are visible everywhere, moss invades and covers parts of the hylaeosaur’s flanks (not good if you want the models to persist) and sections of the megalosaur’s nose look like they could fall off at any moment. Similar damage is present on some of the other models, their skin and scales flaking or cracking or looking to be in imminent danger of breaking or falling off. Some substantial (expensive) repair work has been done by the Friends of Crystal Palace Dinosaurs, but much more is required.

Caption: looking into the mouth of the Mosasaurus. “The large pointed teeth on the jaws are very conspicuous; but, in addition to these, the gigantic reptile had teeth on a bone of the roof of the mouth (the pterygoid), like some of the modern lizards” (Owen 2013, p. 11). Image: Darren Naish.
In my previous Crystal Palace article, I discussed the fact that the models reveal a great many complex anatomical details, some of them involving details only familiar to specialists. When you see the models up close, even more such details become apparent. I’m not sure I knew that the mosasaur is equipped with accurate palatal teeth, for example. Owen specifically referred to this feature in the guidebook (Owen 2013). The temnospondyls (‘labyrinthodonts’) have big palatal teeth as well, as they should.

Caption: anterior view of one of the temnospondyls. Check out the accurate palatal teeth. To Owen and Hawkins, this animal was Labyrinthodon salamandroides, a sort of composite based on temnospondyl bones and teeth, and inferences made from croc-line archosaur footprints, thought by Owen and those who followed his work to be made by Labyrinthodon. Image: Darren Naish.
Like the dinosaurs (except the megalosaur), the surviving pterosaurs are fantastically scaly (today, we think that pterosaurs were covered in a filamentous coat, except on their wings, the distal parts of their hindlimbs and their snouts and faces). Unfortunately, the pterosaur with folded wings has recently been damaged, its smashed snout and lower jaw meaning that you can see right inside its head. This reveals a complex internal ‘anatomy’: another reminder that the models weren’t all built to the same plan or in the same style, but that very different approaches were used for each.
Caption: the two large Crystal Palace pterosaurs represent the species known to Owen and Hawkins as Pterodactylus cuvieri (though the possibility that more than one species is represented is raised by Owen’s remarks in the accompanying guide). Unfortunately, one of the models is now badly broken. The two smaller pterosaur models are not currently on display and have had a really unfortunate history: they’ve been vandalised, broken and stolen several times. Image: Darren Naish.
Both big pterosaurs stand atop a small rocky ’cliff’. Like all the geological structures in the park, this is an installation specially created as part of the display. It looks, at first sight, to be made of nondescript grey rock. While looking at it, I began wondering about its specific composition, since we know that the other chunks of rocks in the park aren’t just random lumps of local geology, but transplanted sections of the specific geological unit the respective animal’s fossils come from.

Caption: as per usual, the model up-close - this is the Pterodactylus cuvieri posed with open wings - is a remarkable bit of craftmanship. Pterodactylus cuvieri was named for bones that have more recently been included within the genera Ornithocheirus and Anhanguera, and have most recently been awarded the new name Cimoliopterus. Image: Darren Naish.

Caption: and here’s a close-up of that detail. I absolutely adore the work here; check out all those individual scales. It seems remarkable now to think that Owen and Hawkins really imagined pterosaurs to look like this, but here’s the evidence. Image: Darren Naish.
What, then, are these pterosaurs really standing on? Mark Witton and I examined some freshly broken fragments of the cliff – the rock is chalk! This really shouldn’t have been a surprise given that the fossils these reconstructions are based on come from the English Chalk (Owen 2013), but it was great to see it confirmed. There’s even a line of dark flint nodules, just as there is in real chalk cliffs. These details are surely known to specialist researchers but were news to me.

Caption: broken sections of the ‘pterosaur cliff’ reveal that we’re looking at chalk… which isn’t a surprise, and is exactly what we would expect, but here’s confirmation. You should be able to see a few of the dark, shiny flint nodules too. Image: Darren Naish.
Look – below – at the photo of the teleosaurs. Notice how the arrangement of scales and scutes is highly detailed, and how the animals have been given a scute arrangement that very much resembles that of living crocodylians. As it happens, the arrangement they’ve been given is dead wrong for teleosaurs but it is absolutely accurate for living crocodylians (where dorsal scute arrangement is – mostly – diagnostic to species level). What I’m saying is that I think that Hawkins looked at living Saltwater crocodiles Crocodylus porosus when designing these amazing models, since their dorsal scute pattern specifically matches this species (and, surprisingly, not gharials).

Caption: the two Teleosaurus of Crystal Palace. While compared by Owen with gharials, it’s interesting that the dorsal scute pattern they were given is very clearly based on living crocodiles. As per usual, look at the remarkable amount of well-rendered detail. Image: Darren Naish.
As usual, there’s stacks more I want to say, but time is up. I had such a great time seeing the models up close and I can’t wait to do it again. At the risk of sounding like a broken record, the Crystal Palace prehistoric animals are among the most scientifically and historically important renditions of ancient creatures ever created, and they’re amazing pieces of art, construction and craftmanship to boot. A full, thorough discussion of their ‘anatomy’, backstory, construction and history has, even today, never been published – McCarthy & Gilbert (1994) is the closest thing to it – and much remains to be compiled and discovered.
These models must be preserved for the future. On that note, don’t forget to pledge your support for the bridge project. Crystal Palace and its models will be covered here again at some point in the future, and various relevant projects will be discussed here in 2019 – watch this space!


As should be obvious from these photos, the entire area has become somewhat overgrown recently, and much maintenance is needed. The Friends of Crystal Palace Dinosaurs group are doing what they can, but help is needed. Image: Darren Naish.
For other TetZoo articles on the Crystal Palace prehistoric animals and other relevant issues, see…
The Great Dinosaur Art Event of 2012, November 2012
All Yesterdays: Unique and Speculative Views of Dinosaurs and Other Prehistoric Animals - the book and the launch event, December 2012
Artistic Depictions of Dinosaurs Have Undergone Two Revolutions, September 2014
The Dinosaurs of Crystal Palace: Among the Most Accurate Renditions of Prehistoric Life Ever Made, August 2016
Refs - -
McCarthy, S. & Gilbert, M. 1994. Crystal Palace Dinosaurs: The Story of the World’s First Prehistoric Sculptures. Crystal Palace Foundation, London.
Naish, D. 2010. Pneumaticity, the early years: Wealden Supergroup dinosaurs and the hypothesis of saurischian pneumaticity. In Moody, R. T. J., Buffetaut, E., Naish, D. & Martill, D. M. (eds) Dinosaurs and Other Extinct Saurians: A Historical Perspective. Geological Society, London, Special Publications 343, pp. 229-236.
Owen, R. 1854. Geology and Inhabitants of the Ancient World. Crystal Palace Library and Euston & Evans, London.
Owen, R. 2013. Geology and Inhabitants of the Ancient World. Euston Grove Press, London.
The Second Edition of Naish and Barrett’s Dinosaurs: How They Lived and Evolved
Regular readers of this blog should know that 2016 saw the publication of the Natural History Museum book Dinosaurs: How They Lived and Evolved, co-authored by this blog’s humble overlord… that might be an oxymoron… and the Natural History Museum’s Paul Barrett (Naish & Barrett 2016). Dinosaurs has been well received and pretty successful in terms of sales, and so it came to pass that there was the need for a modified, softback version that included updates and corrections. Officially, the new version is a ‘fully revised and updated’ version, but it’s very literally a second edition, and that’s what I’m calling it.

Naish and Barrett, second edition - with a new cover!
First off, the production of a second edition – we’ll call it Dinosaurs 2nd ed (Naish & Barrett 2018) from hereon – allowed the correction of assorted typos and poor word choices. It never ceases to amaze me how much stuff we miss even when a given piece of text is checked, double-checked and checked again. Big thanks to fan of the book Klinsman Hinjaya for noting a number of required corrections. More importantly, said second edition also allowed us to update or modify various aspects of the science covered in the book. New data and new interpretations mean that our ideas on the biology and evolutionary history of extinct animals are constantly changing, and this was a great opportunity to get some of the relevant changes incorporated.
Some of you – especially those in possession of the first edition – are keen to know what’s different about Dinosaurs 2nd ed, so – without giving too much away – let’s take a look…

Caption: some of Emma’s drawings feature in the book. No, I’m kidding - they don’t. Or do they. Credit: Darren Naish.
A new cover. Personally, I think that Dinosaurs: How They Lived and Evolved is a pretty good book, and I hope you agree. But I never much liked the cover, and I know I’m not alone. Given that it portrays a roaring monster that’s showing us the inside of its mouth and what biiiig teeth it has, it might be construed as being contrary to the message otherwise promoted throughout the book: that non-bird dinosaurs were animals, and not the monsters of Hollywood and popular fiction.

Caption: new cover art by Bob Nicholls of Paleocreations. I own a full-sized print of this amazing piece. Damn… I own a lot of Bob Nicholls art now. Credit: Bob Nicholls.
The plan for Dinosaurs 2nd ed was thus to produce a brand-new cover that better represented modern thinking on dinosaurs. Paul and I opted to have either a feathered theropod, or an unusual ornithischian, and we ended up going with the latter for reasons I can’t recall (I think because it might look weirder and less familiar: feathered theropods are so passé, after all). Our chosen artist – Bob Nicholls – came up with a bunch of test sketches, all depicting the Chinese heterodontosaur Tianyulong in various poses and behavioural settings. We chose one, and Bob went to extraordinary trouble to get the contrast, lighting and composition right. There’s much more to it than that but… ladies and gentlemen, I give you… our new cover.
New artwork. Moving now to the insides of the book, we’ve also replaced a few other images that were used in the first edition. Bob has a few more of his excellent images in the book, we swapped out the (now defunct) diagrams on diplodocid jaw movement with a new reconstruction of a ground-feeding diplodocid (though illustrated without the keratinous beak recently proposed for this group), and Matt Martyniuk’s Anchiornis replaces a previous image with a problematic forelimb configuration. I also replaced a bunch of small images used in various of the cladograms.
Caption: some of the cladograms of Dinosaurs 2nd ed have been tweaked a little. This one depicts Theropoda, the predatory dinosaurs. Credit: Darren Naish.
Necessary coverage of the Ornithoscelida thing. Currently, one of the most talked-about issues within the study of Mesozoic dinosaurs is Matt Baron et al.’s (2017) proposal that the main branches of the dinosaur family tree be rearranged, such that sauropodomorphs are outside a theropod + ornithischian clade termed Ornithoscelida. I wrote about this proposal when it was brand-new, back in March 2017. It has been hotly contested by several teams of authors and there are already a few publications saying how reanalysis does not support Baron et al.’s model, or at least does not support it preferentially above the others that are available.
Obviously, we just had to cover this issue for Dinosaurs 2nd ed – especially so given that Paul is one of the study’s authors – and a few new paragraphs of text and new diagrams summarise the area for our readers. Our coverage of the Ornithoscelida issue resulted in various knock-on changes elsewhere in the book. New phylogenetic positions, for example, have been favoured for herrerasaurids, Eoraptor and so on.
Caption: the Baron et al. model, as depicted in the 2017 article I wrote about it for TetZoo ver 2. Credit: Darren Naish.
New taxa, new names, new phylogenetic possibilities. The world of Mesozoic dinosaur phylogeny and systematics move fast – remember that an average year right now sees the naming of between 30 and 50 new non-bird dinosaur species. We’re at a stage where phylogenetic models are never really ‘overturned’, but they certainly undergo regular tweaking, modification and augmentation. In view of this, Dinosaurs 2nd ed includes references to Stenonychosaurus, Latenivenatrix (sorry, Troodon) and halszkaraptorines, and I subtly changed the wording on the megaraptorans…
Pisanosaurus is no longer bigged-up as a possible early ornithischian given data indicating that it’s a non-dinosaur.
Revising thoughts on the origins of flight. Those familiar with discussions on both bird and flight origins within dinosaurs will know all about the ‘trees down vs ground up’ argument, and also with the contention that it might be utterly wrong to polarise things in this way. Nevertheless, there remain – for all those attempts to point to shades of grey – extremes in the debate. My take on the earliest phases of maniraptoran flight has mostly involved a weird sort of hybrid whereby the animals concerned are deemed predominantly terrestrial but also capable of climbing, their leaping, fluttering and gliding in arboreal settings being antecedent to flight. Like many people, I was originally enthused enough by WAIR (wing-assisted incline running) to think that it might be a plausible explanation of how maniraptorans first came to exploit arboreal settings and, from there, evolve flight.

Caption: Dececchi et al. (2016) showed that at least some non-bird maniraptorans do not have the right combination of anatomical features to benefit from WAIR as originally envisioned. This work affected our thinking on flight origins in Dinosaurs 2nd ed. Credit: Dececchi et al. 2016, PeerJ
The diversity of non-bird maniraptorans is such that it looks likely that these animals practised all sorts of behaviours during the long time that they were around, and thus that various different acts and adventures could have contributed to their ability to leave the ground. Having said that, recent studies indicate that at least some of the relevant animals could likely leap into flight from a ground-based start (Dececchi et al. 2016), and – at the same time – that arboreal behaviour was unlikely in such species. The possibility that flight could well have evolved without any arboreal component is interesting (and even shocking to some), and sufficiently so that we’ve alluded to it (briefly) in Dinosaurs 2nd ed.

Caption: Dinosaurs, the Russian edition. Now I know what my name looks like in Russian. Yes, the title is not the same as the English one.
And that’s it. I should also add that Dinosaurs: How They Lived and Evolved has also recently appeared in Australia (where it’s published by CSIRO), and that a Russian translation is now out as well. My thanks to everyone who’s bought this book, to those who commented on or said nice positive things about the first edition, and to everyone involved in its production and publication.
For those who haven’t purchased a copy, it’s available here from amazon, here from amazon.co.uk, and here from the publishers.
For previous articles relevant to this one, see…
Flight of the Microraptor, November 2013
The Integrated Maniraptoran, Part 1, May 2016
The Integrated Maniraptoran, Part 2: Meet the Maniraptorans, May 2016
The Integrated Maniraptoran, Part 3: Feathers Did Not Evolve in an Aerodynamic Context, May 2016
Naish and Barrett's Dinosaurs: How They Lived and Evolved, November 2016
Ornithoscelida Rises: A New Family Tree for Dinosaurs, March 2017
Refs - -
Baron, M. G., Norman, D. B. & Barrett, P. M. 2017. A new hypothesis of dinosaur relationships and early dinosaur evolution. Nature 543, 501-506.
Naish, D. & Barrett, P. M. 2016. Dinosaurs: How They Lived and Evolved. The Natural History Museum, London.



