I have elected to republish my articles on caecilians. (1) Because TetZoo always needs more amphibian coverage. (2) Because caecilians are awesome. The text you’re about to read – assuming you choose to continue – first appeared in 2008 (original here); I haven’t much updated it. Here we go…

Caption: the flattened and very striking head of the large typhlonectid caecilian Atretochoana, from Wilkinson & Nussbaum (1997).
If I told you that there was a group of living tetrapods that have sensory tentacles, sometimes have protrusible eyes, sometimes lack eyes entirely, often exhibit sophisticated parental care and may even feed their babies on a specially grown layer of nutritious epidermal skin, are incredibly long-bodied yet often lack tails, and sometimes possess large, anatomically complex, eversible male sexual organs, you might wonder which recreational drugs I was taking. The fact is, I'm talking about a very real – albeit very poorly known – group of living tetrapods: the caecilians.

Caption: the caeciliid Caecilia subnigricans of Colombia and Venezuela, demonstrating the worm-like demeanour of this group. Image: José Gabriel Julio Guzmán, CC BY 4.0 (original here).
All extant caecilians are long-bodied, limbless, superficially worm-like amphibians with reduced eyes. They are predominantly fossorial (adapted for burrowing), although some are aquatic or semiaquatic and some terrestrial species have aquatic larvae. Limb girdles are entirely absent and skin folds (annulae) encircle the body.
Despite their mostly fossorial habits some species are visually striking, being blue, purple, green, orange, yellow, or with bright yellow lateral stripes. Others are decorated with blotches, or have a head that’s lighter in colour than the body. Poison glands are present (some species may even be quite toxic to humans), so it's likely that the bright colours are aposematic. While the smallest species (like the grandisoniid Idiocranium russelli from Cameroon) are mature at just 70 mm, the largest – the very slender-bodied Caecilia thompsoni from Colombia – reaches 1.5 m.
Caption: the bright orange São Tomé caecilian Schistometopum thomense, a dermophiid from islands in the Gulf of Guinea (reports of a similar animal from the mainland might represent an additional species). Image: Marius Burger, CC0 (original here).
Food, feeding and predation. In what appear to be the most anatomically archaic of living caecilians (the South American rhinatrematids), a short tail is present and the mouth is located at the front of the head; in the more advanced groups, no tail is present and the mouth is situated further back, slung underneath the head. It's mostly assumed that caecilians are generalist predators of soil invertebrates that feed opportunistically on earthworms, ants and termites: animals that have been termed 'soil ecosystem engineers' by ecologists.
However, it's also been argued that some caecilians are specialists (preying on specific termite or earthworm species, or on beetle pupae), or even detritivores, ingesting leaf humus and other plant fragments (Hebrard et al. 1992). This last proposal is radical given that amphibians are essentially all (with the exception of a handful of weirdos) carnivorous as adults, and a later study showed that soil and detritus recovered from the gut contents of the species concerned (the east African herpelid Boulengerula taitana) originated from its earthworm prey (Gaborieau & Measey 2004). Small vertebrates including frogs, lizards, burrowing snakes (Presswell et al. 2002) and possibly rodents sometimes fall prey to large caecilians. Caecilians themselves are preyed upon by burrowing snakes, fishes (Gazola Da Silva et al. 2007), and by chickens, pigs and the tenrecs that have been introduced to the Seychelles.

Caption: excellent illustration of how confident a caecilian might be about its predatory prowess. This photo was taken in the Rio Zunac Reserve, Ecuador, by reserve guards Luis and Fausto Recalde. It is from here at the Fundacion EcoMinga blog.
Distribution, ecology, taxonomy. Caecilians occur throughout the humid tropical regions of the world: in South America, equatorial eastern and western Africa, and tropical Asia including the Philippines and the western Indo-Australian archipelago. They also inhabit the Seychelles. The absence of caecilians from central equatorial Africa and Madagascar is odd and we should consider the possibility that they might await discovery in these areas. Generally assumed to be rare and highly elusive, some studies have shown that at least some caecilian species can be abundant if searched for (Measey 2004), and in fact present in sufficient numbers to be (presumably) ecologically significant, especially so given their effects on soil ecosystem engineers (Jones et al. 2006).
It stands to reason that, because of their fossoriality, we know relatively little about caecilians, and because many species are poorly known and poorly represented in zoological collections, confusion about their systematics and taxonomy has been rife. A nice example is provided by the tropical American dermophiid Gymnopis* syntrema: in a paper subtitled ‘a comedy of errors’, Nussbaum (1988) argued that “six different binomial names have been applied to this species; it has been assigned to seven genera, two subfamilies of a single family, and three families” (p. 921).
* Not a typo, though of course some workers have inadvertently spelt it Gymnophis [sic].

Caption: this article doesn’t discuss the aquatic typhlonectid caecilians, which are – ironically – the only caecilians the majority of us non-specialists get to encounter. Here’s a captive Typhlonectes natans eating tubifex worms. Image: Neil Phillips, used with permission.
How many taxa, how many families? Currently (as of October 2022), 33 genera and 215 caecilian species are recognised, so this isn't a huge group. The 200th species was named in 2015; there was much rejoicing, and a special song was written.
Until 1968 all caecilians were lumped into a single taxonomic family – dubbed Caeciliidae** – but later work by Edward H. Taylor (read on) and Ronald A. Nussbaum proposed that this should be divided into six families (Rhinatrematidae, Ichthyophiidae, Uraeotyphlidae, Scolecomorphidae, Caeciliidae and Typhlonectidae). Uraeotyphlids are a tropical Asian group, and it’s now agreed that they should be considered synonymous with the Asian scaled caecilians, or ichthyophiids.
** Little known is that a family of psocodean insects has also been called Caeciliidae, and it was recommended by the ICZN that the caecilian group therefore go by the older spelling Caeciliaidae (psocodeans are booklice, parasitic lice and so on). Nussbaum & Wilkinson (1989) argued that this decision was inappropriate because Rafinesque, the author of Caeciliaidae, never associated any generic name with this group (he only named it for the Italian vernacular word 'Ceciliani'), nor did he specify its intended content. Despite these objections, Nussbaum & Wilkinson (1989) continued to use Caeciliaidae following the ICZN's ruling, although other authors did not. In 1996 the ICZN changed the ruling such that Caeciliidae became reinstated for the amphibians; the alternative name Caeciliusidae is now used for the psocodeans (Wilkinson & Nussbaum 2006).

Caption: at left, holotype of the Indian caecilian Chikila darlong. Scale bars are 20 mm in A, 10 mm in B-G. At right, a female Herpele squalostoma from Cameroon with young. These two animals represent two caecilian family-level lineages only recently named or resurrected from synonomy, namely Chikilidae and Herpelidae. Images: Chikila from Kamei et al. (2013), CC0; Herpele from Kouete et al. (2012), CC BY 3.0.
Recent discoveries have upped the number of recognised caecilian families to ten. The five additional families are the Neotropical and African group Dermophiidae (named in 1969, then sunk, then resurrected by Wilkinson et al. (2011)), the north-east Indian caecilians or Chikilidae (named for the south Asian Chikila, published in 2012), the African caecilians or Herpelidae (named in 1984, then sunk, but resurrected by Wilkinson et al. (2011)), the Indo-African caecilians or Grandisoniidae (named in 1986, then sunk, but also resurrected by Wilkinson et al. (2011), and the South American caecilians or Siphonopidae (named in 1850, then sunk, then also resurrected by Wilkinson et al. (2011)). Most studies find the sort of phylogeny depicted below (from Kamei et al. (2012)).

Caption: a phylogenetic hypothesis for caecilians, from Kamei et al. (2012). The American tailed rhinatrematids and Asian ichthyophiids are the earliest-diverging members of the group. This cladogram obviously emphasises the position of the recently recognised chikilids.
Books on caecilians. There are very few good non-technical sources where you can read up on caecilians: the only one I can readily recommend is Nussbaum (2000). A major volume devoted entirely to caecilians, Edward H. Taylor's 1968 848-page-long The Caecilians of the World: A Taxonomic Review is cited in just about every bit of caecilian literature and proved instrumental in initiating new interest in the taxonomy and phylogeny of the group. I thought about getting hold of a copy, but gave up when I discovered that it can't be obtained for less than £138 ($275) [UPDATE: those are 2008 prices. Today it’s available for sale at £305/$350]. A German book, Die Blindwühlen, was published on the group in 1996 (Himstedt 1996), and a French one – Les Gymnophiones, ces curieux Amphibiens – in 2000 (Exbrayat 2000).

Caption: the sort of book that really should belong in the Tet Zoo Towers library, hint hint. Yes, images of Taylor’s 1968 The Caecilians of the World. These images from here at AbeBooks.
Tentacles and protrusible eyes. In some details, caecilian anatomy is surreal. Easily the weirdest features have to be the tentacles. Unique to the group, these are short paired sensory structures that emerge from a cavity on the side of the snout between the eye and nostril. They are always quite easy to see, even in the smallest species. No other tetrapod has anything like this. Derived from the tear duct, extrinsic eye muscles and other orbital structures, the tentacles are connected to the vomeronasal organs and presumably allow the animals to test their environment for sensory clues.

Caption: the robust, bullet-shaped skull of the American dermophiid Dermophis mexicanus. This specimen was collected in Guatemala and made available for scanning by David Cannatella and the Amphibian Tree of Life project. It was scanned for DigiMorph; other images and files are available here.
The caecilian skull is generally bullet-shaped, robust, thick-boned and with strongly adhering skin. Their recurved teeth are sometimes sharp-keeled and bicusped. The eyes of caecilians are sometimes visible beneath the skin and set within bony sockets (e.g., Ichthyophis), are sometimes hidden beneath the bones of the skull (e.g., Scolecomorphus), and are sometimes completely absent (e.g., Boulengerula). While the tentacle is often located close to the nostril and some distance from the eye, the eye and tentacle are close in position in some species.
Scolecomorphids, containing only Scolecomorphus and Crotaphatrema, are unique to equatorial Africa and have a particularly large tentacular opening located near the tip of the snout, ahead of the under-slung mouth. The close position of the eye and tentacle mean that they've become connected: in its resting position, the eye is located beneath the lateral surface of the skull, but full extrusion of the tentacle causes the eye to move out of the skull and down the tentacle (O'Reilly et al. 1996). An area of the tentacle lacking in pigmentation presumably allows light to reach the retina. Scolecomorphids are the only tetrapods that can deliberately move their eyes out of their skulls. Scolecomorphids are also unusual in lacking a stapes.

Caption: close-ups of Scolecomorphus kirkii, showing how the eye is mobile and can be moved from the tentacular groove on the side of the face (a) and then into the tentacle when it’s extended (b). These images are from O'Reilly et al. (1996).
Caecilians are also unique in that they have two sets of jaw-closing muscles: in addition to the adductor mandibulae muscles that are located at the back of the jaws, they also use their interhyoideus posterior muscles in jaw closure. These muscles are located in the neck and are attached to the retroarticular process at the posterior end of the lower jaw: they close it by pulling down and backwards on the retroarticular process (Nussbaum 1983). Other tetrapods possess interhyoideus posterior muscles, but they aren't used in jaw closing, so why have caecilians co-opted them for this use? The answer might be that, in evolving a rigid, box-like skull specialised for burrowing, caecilians have had to reduce and close the temporal fossae that originally housed the adductor mandibulae muscles. To compensate for the weakened power provided by the now reduced adductor mandibulae, caecilians have switched to using the interhyoideus posterior as well. This looks like a classic example of exaptation, the process whereby an existing structure is co-opted for a new function.
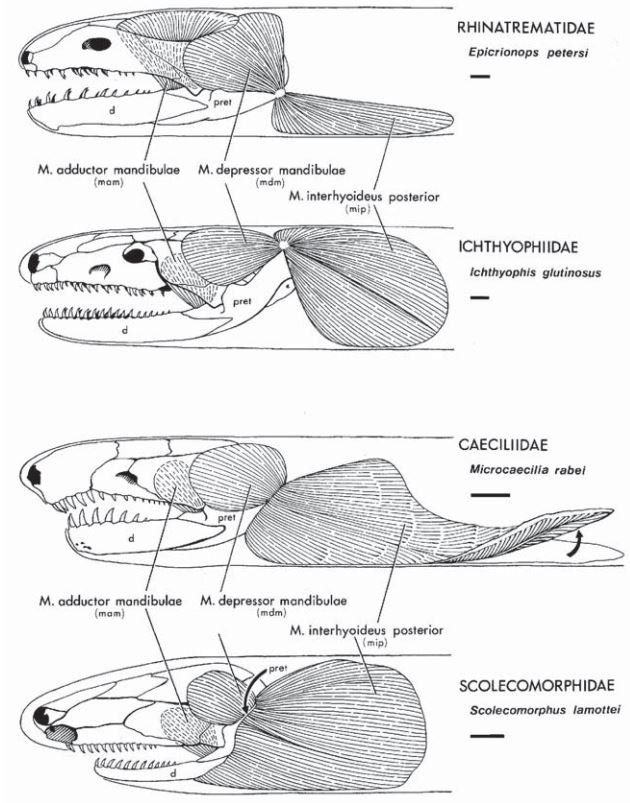
Caption: variation in caecilian cranial and interhyoideus posterior morphology, as illustrated by Wilkinson & Nussbaum (2006).
Of course, this isn't where it ends: we still have the weird sexual organs to look at yet, plus the stegokrotaphy, zygokrotaphy, the whole 'out of India' thing, the viviparity, dermatotrophy and matrotrophy, and the biggest lungless tetrapod ever (the bizarre Atretochoana, shown at the very top of the article). Be sure to tune in to the next thrilling instalment of Tetrapod Zoology!
For previous articles on caecilians and other amphibians, see…
Surreal caecilians part I: tentacles and protrusible eyes, January 2008
Surreal caecilians part II: pass mum’s skin, hold the mayo, January 2008
‘The Secret World of Naked Snakes’: a ZSL event, November 2009
Carnivorous, worm-like amphibians invade London: ‘The Secret World of Naked Snakes’, part I, December 2009
Yummy mummies, caecilians on the EDGE, and the gigantic Minhocão: ‘The Secret World of Naked Snakes’, part II, December 2009
Because caecilians are important, August 2013
Refs - -
Exbrayat, J.-M. 2000. Les Gymnophiones, ces curieux Amphibiens. Edition Boubée, Paris.
Frost, D. R., Grant, T., Faivovich, J., Bain, R. H., Haas, A., Haddad, C. F. B., De Sá, R. O., Channing, A., Wilkinson, M., Donnellan, S. C., Raxworthy, C. J., Campbell, J. A., Blotto, B. L., Moler, P., Drewes, R. C., Nussbaum, R. A., Lynch, J. D., Green, D. M. & Wheeler, W. C. 2006. The amphibian tree of life. Bulletin of the American Museum of Natural History 297, 1-370.
Gaborieau, O. & Measey, G. J. 2004. Termitivore or detritivore? A quantitative investigation into the diet of the east African caecilian Boulengerula taitanus (Amphibia: Gymnophiona: Caeciliidae). Animal Biology 54, 45-56.
Gazola Da Silva, F. F., Mott, T., Garey, M. V., & Vutule, J. R. S. 2007. Chthonerpeton viviparum Parker & Wettstein, 1929 (Amphibia, Gymnophiona, Typhlonectidae) in Paraná state, Brazil and the first record of predation of this species by Hoplias malabaricus (Bloch, 1794) (Actinopterygii, Erythrinidae). Pan-American Journal of Aquatic Science 2, 261-262.
Hebrard, J., Maloiy, G. & Alliangana, D. 1992. Notes on the habitat and diet of Afrocaecilia taitana (Amphibia, Gymnophiona). Journal of Herpetology 26, 513-515.
Himstedt, W. 1996. Die Blindwühlen. Westarp, Magdeburg.
Jones, D. T., Loader, S. P. & Gower, D. J. 2006. Trophic ecology of east African caecilians (Amphibia: Gymnophiona), and their impact on forest soil invertebrates. Journal of Zoology 268, 117-126.
Kamei, R. G., Gower, D. J., Wilkinson, M. & Biju, S. D. 2013. Systematics of the caecilian family Chikilidae (Amphibia: Gymnophiona) with the description of three new species of Chikila from northeast India. Zootaxa 3666, 401-435.
Kamei, R. G., San Mauro, D., Gower, D. J., Van Bocxlaer, I., Sherratt, E., Thomas, A., Babu, S., Bossuyt, F., Wilkinson, M. & Biju, S. D. 2012. Discovery of a new family of amphibians from northeast India with ancient links to Africa. Proceedings of the Royal Society B 279, 2396-2401.
Measey, G. J. 2004. Are caecilians rare? An east African perspective. Journal of East African Natural History 93, 1-21.
Nussbaum, R. A. 1983. The evolution of a unique dual jaw-closing mechanism in caecilians (Amphibia: Gymnophiona) and its bearing on caecilian ancestry. Journal of Zoology 199, 545-554.
Nussbaum, R. A. 1988. On the status of Copeotyphlinus syntremus, Gymnopis oligozona, and Minascaecilia sartoria (Gymnophiona, Caeciliidae): a comedy of errors. Copeia 1988, 921-928.
Nussbaum, R. A. 2000. Caecilians. In Cogger, H. G., Gould, E., Forshaw, J., McKay, G. & Zweifel, R. G. (consultant eds) Encyclopedia of Animals: Mammals, Birds, Reptiles, Amphibians. Fog City Press (San Francisco), pp. 492-499.
Nussbaum, R. A. & Wilkinson, M. 1989. On the classification and phylogeny of caecilians (Amphibia: Gymnophiona), a critical review. Herpetological Monographs 3, 1-42.
O'Reilly, J. C., Nussbaum, R. A. & Boone, D. 1996. Vertebrate with protrusible eyes. Nature 382, 33.
Presswell, B., Gower, D. J., Oommen, O. V., Measey, G. J. & Wilkinson, M. 2002. Scolecophidian snakes in the diets of south Asian caecilian amphibians. Herpetological Journal 12, 123-126.
Wilkinson, M. & Nussbaum, R. A. 1997. Comparative morphology and evolution of the lungless caecilian Atretochoana eiselti (Taylor) (Amphibia: Gymnophiona: Typhlonectidae). Biological Journal of the Linnean Society 62, 39-109.
Wilkinson, M. & Nussbaum, R. A. 2006. Caecilian phylogeny and classification. In Exbrayat, J.-M. (ed.) Reproductive Biology and Phylogeny of Amphibia Volume 3. Gymnophiona. Science Publishers Inc., pp. 39-78.



