In 1961, biologist Harald Stümpke published his famous work Bau und Leben der Rhinogradentia….

Caption: at left, my own copy of Stümpke (1961), which I own thanks to the good graces of Dr Mathew Wedel. At right: a copy posed with an Earwing skeleton (photo by Dr Michael Granatosky).
Therein, he described the recently discovered, island-dwelling mammal group known vernacularly as the snouters or rhinogrades and technically as the rhinogradentians. The snouters exhibited unprecedented diversity in form, ecology and habits. While originating from shrew-like ancestors, they included volant, subterranean, sessile, predatory, herbivorous and even miniaturised, parasitic forms, and I use the past tense because the group seemingly became extinct due to an atomic accident (though read on). Such is snouter diversity that they’ve remained a classic example of evolutionary diversification and island endemism in mammals: numerous post-1961 papers, textbooks and articles discuss their biology and evolution, while continued interest in the group is perpetuated by the documentation of new fossil and extant species, some of which have been reported from continental and marine settings (Feldoyants & Zelbstander 2000, Bukashkina 2004, Kashkina 2004).
Such is snouter importance in mammalogical literature that I couldn’t ignore them here at Tetrapod Zoology, and in fact I published a lengthy, two-part review of the group early in the blog’s history, back in 2007 (they’re here and here, should you wish to check them for historical reasons). Because those articles are – in intact form – now only findable at wayback machine, it seemed right to republish them today, and here we are. For those of you learning about snouters for the first time, get ready for a wild ride….

Caption: Stümpke’s book includes a huge amount of insight on the evolutionary history and phylogenetic relationships of these animals, and also makes it clear that many issues were unresolved and warranted further study. At left is the phyllogram included in the book (actually, this version – borrowed from a website – is different in some respects from the version present in the edition I have); at right is my effort to depict what it shows more schematically. Illustrations from Stümpke (1961).
Ok, at this point I’ll break character and tell you that the snouters never were real, and nor was Harald Stümpke. Stümpke was a pseudonym of German zoologist Gerolf Karl Steiner (1908–2009) who invented the snouters, and their entire backstory and geographical setting, while teaching at Heidelberg and Darmstadt during the 1940s. It’s somewhat unclear whether the book which eventually resulted – translated into English in the USA in 1967 – was intended as a sort of sophisticated in-joke among zoologists and perhaps scientists more widely, or created to be partly instructional, and to highlight the themes, terminology and theories of biology. An essay exploring these ideas, and providing much useful biographical information on Steiner and the snouter backstory, has recently been published by Cain (2018). I should add, incidentally, that I disagree with Cain’s main premise (that the popularity and persistence of snouters in literature is due to ingroup/outgroup tribalism): his approach, in my opinion, misses the reason why so many of us enjoy, and engage with and popularise, speculative biology.
I consider myself very lucky to have obtained Stümpke’s book when I did (mine is a 1967 Natural History Press edition, shipped from the Church College of Hawaii), since it has accrued in value in recent years and is now only available at astronomical, prohibitive prices.
With the above in mind, find below my snouter-themed writings as originally used at TetZoo ver 2 back in 2007. This text was published as two articles but I’m here combining them into one. I haven’t really updated the text except for style and to make reference to post-2007 discoveries. I have been able to add much better images, partly because better ones are now available but also because I’m more flexible in image creation now than I was in 2007. Anyway, to business…
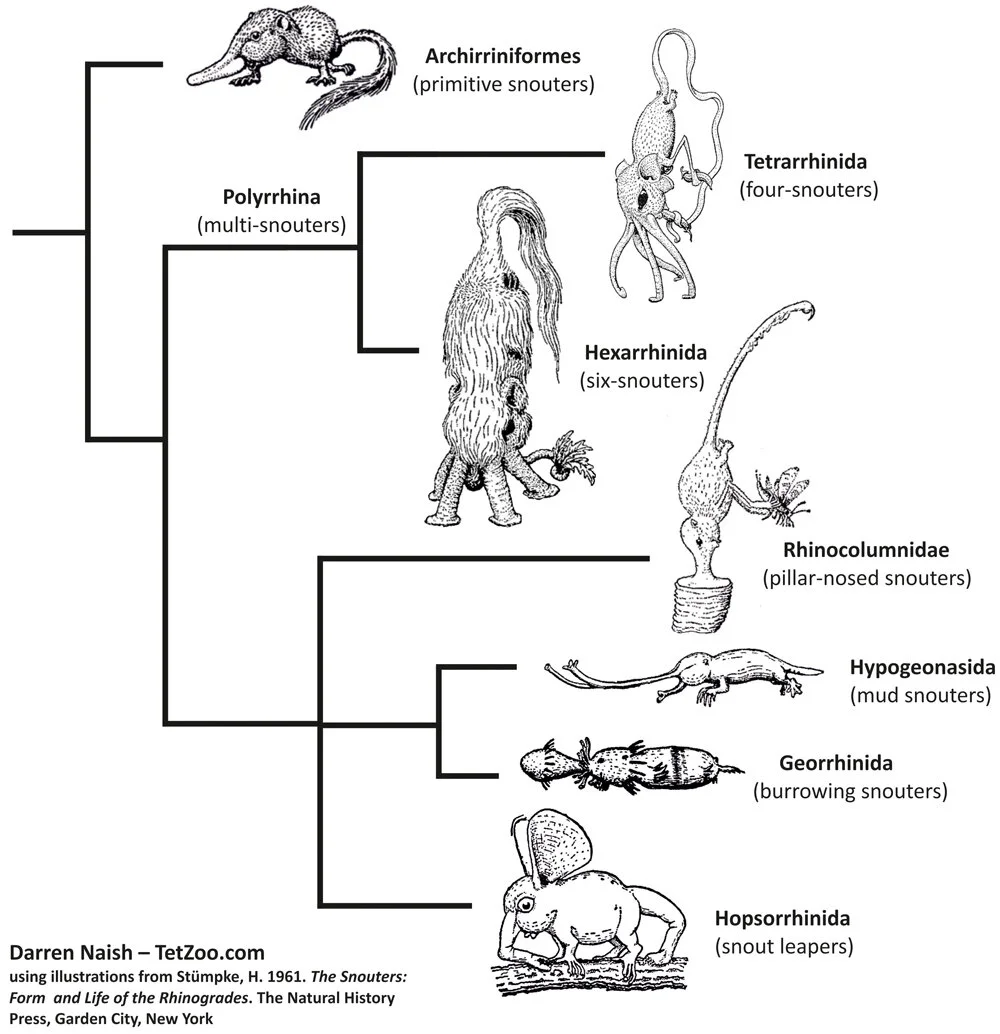
Caption: an effort to depict rhinogradentian phylogeny, based on the perceived affinities and characters discussed by Stümpke (1961). It appears that all post-archirrhiniform snouters form a clade which diverged early on into two. Stümpke’s ‘monorrhinan’ group is an artificial group according to this view. After completing this diagram I discovered that other efforts to depict snouter phylogeny exist: another can be observed here. Illustrations from Stümpke (1961).
Introducing the snouters. I’m always surprised when I meet zoologists who aren’t familiar with Harald Stümpke’s famous 1961 book Bau und Leben der Rhinogradentia, a volume translated into English by Leigh Chadwick in 1967 as The Snouters: Form and Life of the Rhinogrades (and referred to from hereon as ‘Stümpke 1961′). For the few who don’t know, this legendary text discusses in marvellous detail the biology, lifestyle and evolution of the snouters (aka rhinogrades or rhinogradentians), a bizarre and unique group of small mammals originally thought endemic to the south Pacific Hi-yi-yi archipelago, an island group made known to westerners by Einar Pettersson-Skämtkvist in 1941 as he escaped from Japanese imprisonment.
Little known is that Skamtkvist didn’t only learn of the rhinogradentians: the islands were also inhabited by the Hooakha-Hutchi people, the bizarre megaphone birds, and the endemic shrew Limnogaloides. A diverse endemic insect fauna was bizarre in including Palaeozoic relicts, and the flora also included archaic forms such as large clubmosses. Of great interest is that rhinogradentians were mentioned in late 19th century European literature, and indeed there is some indication that the Hi-yi-yi islands had actually been discovered by westerners long prior to 1941 (Stümpke 1961).

Caption: a snouter montage, depicting the better part of anatomical diversity within the group. All of the animals shown here are discussed and (mostly) illustrated further below. Illustrations from Stümpke (1961).
As evidenced by the bibliography of Stümpke (1961), a substantial number of publications devoted to rhinogradentian morphology, behaviour and evolution appeared during the 1940s and 50s and such was the interest in this group that the Darwin Institute was set up on Hy-dud-dye-fee (the largest island of the archipelago), and an international conference on rhinogradentians held there in 1953. In what must be ranked as one of the greatest human-made environmental catastrophes of all time, secretly conducted atomic tests occurring in the same region initiated an earthquake that caused the sudden subsidence and destruction of the entire archipelago. Not only were all rhinogradentians known at the time made extinct; all scientists formally specialising on the group were killed. As we’ll see later, rhinogradentians have proved not to be endemic to the Hi-yi-yi archipelago as was assumed, and the discovery of new rhinogradentian species has continued to the present, making the group a constant presence in the zoological literature.

Caption: rhinogradentian experts, gather at the Darwin Institute in Hy-dud-dye-fee in 1953. This photo is taken from here at the excellent French Les Rhinogrades site.
What’s always bugged me about rhinogradentians is that information on many of the taxa that Stümpke described is impossible to come by, unless you’re lucky enough to own a copy of Stümpke (1961). 14 family-level taxa and 189 species were named, and the possibility that additional, undiscovered species might have inhabited the archipelago was regarded as likely. Partly because Stümpke’s book is hard to get hold of in many parts of the world, the full diversity of rhinogradentians is not as well appreciated as it might be. A few internet sites discuss rhinogradentian diversity and figure a few of the rarely-discussed species (most notably, Les Rhinogrades), but they fail to cover the more remarkable lineages within the group*. In order to amend things, and to bring an understanding of rhinogradentian diversity to a greater audience, my aim here is to briefly review the group in entirety. Let me emphasise that I’m barely providing a fraction of the information published within Stümpke (1961), however, and if you want in-depth information on the members of the group you must get hold of the book yourself.
* Since I wrote this text in 2007, things have improved somewhat. The book’s text is reproduced here at Sivatherium and the Speculative Evolution wiki reviews snouter diversity here.

Caption: reconstructions of Archirrhinos, at left from Stümpke (1961); at right, a coloured image produced during the early 2000s. I can find various versions of this image online but not the original. Let me know if you can help as I want to credit the artist.
‘Primitive snouters’, snail-like snouters, sugarmice and others. Archirrhinos haeckelii, Haeckel’s primitive snouter, was identified by Stümpke (1961) as the ‘most archaic’ rhinogradentian: as the one morphologically closest to the ancestral form of the entire group. A superficially shrew-like quadruped, it possessed a large nasarium that served as a support when the animal lunged forward to grab prey. Archirrhinos -like ancestors are thought to have given rise to the nasolimacids, the snail-like snouters. While primitively similar to Archirrhinos, nasolimacids evolved a specialised nasarium where the muscles and sinuses were extensively subdivided, thereby allowing the structure to function much like the foot of a snail. These remarkable mammals lived standing on their foot-like nasarium, their bodies sub-vertical and their limbs reduced or modified. They included armour-plated forms as well as the sugarmice Rhinolimaceus, a taxon so named as a gland at its tail base secreted a sweet-tasting fluid.

Caption: the Snuffling sniffler Emunctator sorbens, a taxon regarded by Stümpke (1961) as a sort of evolutionary connection between snail-like snouters and pillar-nosed snouters. This is my least favourite of the snouters, an opinion probably based on its grotesque lifestyle but mostly its overly human-like face. Images: at left, life-sized model on show at Strasbourg Zoological Museum, photo CC BY-SA 3.0 (original here); at right, from Stümpke (1961).
Probably closely related to the nasolimacids were the rhinocolumnids or pillar-nosed snouters. Again, what was probably the most basal member of the group – the snifflers Emunctator – were not all that different from Archirrhinos, though they differed notably from that taxon in using mucosal strands to trap and ingest prey from shallow water. Emunctator possessed a long, mobile tail equipped with a venomous spur at its tip, and the same character was present in most other pillar-nosed snouters. Among these are the sedentary honeytails Dulcicauda, a group that – like the sugarmice – evolved caudal glands that secreted fragrant secretions that were attractive to insects (Stümpke 1961).
The pillar-nosed snouters of the short-tailed genus Columnifax developed a remarkable symbiosis with various of the predatory snout leapers, of which we’ll see more of later. Columnifax produced milk that the snout leaper drank, while the snout leaper would collect hermit crabs that were eaten by the Columnifax (Stümpke 1961).

Caption: the snail-like nasolimacids (like the slime snouter at left) and pillar-nosed rhinocolumnids (like the honeytail shown at right) appear to have formed a clade for which the name Epigeonasida can be used. Illustrations from Stümpke (1961).
Ribbon snouters and trumpet snouters. What particularly captured the attention of many of those who studied rhinogradentians is that, though descending from shrew-like terrestrial insectivores, they underwent one of the most remarkable adaptive radiations seen among mammals. In addition to the terrestrial forms we’ve seen so far, there were also fossorial, aquatic and arboreal forms. A morphologically unusual and phylogenetically isolated group, the hypogeonasidans, included both fossorial and aquatic forms. Among the fossorial species were the bizarre ribbon snouters Rhinotaenia: a group of small, blind mole-like rhinogradentians with a bilobed, asymmetrical, tube-like siphonal nasarium (unfortunately, the name ‘ribbon snouter’ is also used for an entirely different group that’s part of the Polyrrhina clade; read on). Predominantly animals of tidal sediments, some of them lived almost like parasites within the valves of large bivalves (Stümpke 1961).

Caption: a hypogeonasidan montage. All members of this highly unusual group were small, subterranean or aquatic animals, some adapted for life in tidal mud (or even inside mollusc shells), others for floating in fresh or brackish water. The Rhinostentor species formed great floating rafts of hundreds of individuals. Illustrations from Stümpke (1961).
Hypogeonasidan ribbon snouters are generally regarded as closely related to the aquatic rhinostentorids, or trumpet snouters. These also possessed an elongate siphonal nasarium, but their absence of features associated with subterranean life suggests that they never went through a sediment-dwelling stage and may instead have descended directed from hypogeonasidans that lived at the water’s edge. Indeed several advanced characters, and a unique sort of mucin secretion, were common to the snifflers and the hypogeonasidan ribbon snouters, suggesting that hypogeonasidans evolved from snifflers (Bromeante de Burlas 1952).
Trumpet snouters are among the groups that are usually neglected whenever rhinogradentians are discussed. In these forms the nasarium was a funnel-like filtering apparatus, formed into a terminal rosette fringed with water-repellent hairs. Stiff bristles lined the naked body. Some species hung suspended from the water’s surface while others were pelagic (Stümpke 1961).
Burrowing snouters. Closely related to the hypogeonasidans were the even more remarkable georrhinidans or burrowing snouters, one of two groups in which the snout was sometimes markedly larger than the rest of the body (the other is Dolichoproata, the tasselsnouters). Anatomically archaic members of Georrhinida are best exemplified by the fossorial mole snouters Rhinotalpa. Equipped with strongly reduced limbs as well as wreaths of stiff bristles around the snout and head, Rhinotalpa used inflation and deflation of its nasarium as well as erection and relaxation of the bristle wreaths to move within its burrows (Stümpke 1961). These features were not present in all members of the genus however, and some (e.g., R. angustinasus) exhibited both tiny size and a highly simplified anatomy: besides a short, simple gut, reduced brain, and absent eyes and nares, they apparently relied on cutaneous respiration.

Caption: Rhinotalpa, the ‘least atypical’ of the georrhinidans. The original illustration that features in Stümpke (1961) is quite small, hence explaining the thick lines of the reproduction here. At bottom, we see Takeshi Tokiwa’s excellent model of this taxon. Unfortunately, all versions of this image online are cut off at far right: I haven’t been able to find a complete version of the image.
Georrhinida includes what must be among the most amazing of all tetrapods, the holorrhinids (allsnouters), so structurally modified that even their tetrapod identity has been doubted (actually, this is not the first time that members of a rhinogradentian clade have been wrongly identified as invertebrates). The most specialised allsnouter was the Turbellarian-like dwarfsnouter Remanonasus menorrhinus. Just 2 mm long, this tiny worm-like mammal has been regarded by some as a free-living turbellarian flatworm, and in fact Stulten (1955) argued that turbellarians were highly derived rhinogradentians!

Caption: georrhinidans arranged into a perceived evolutionary sequence (from left to right), showing progressive anatomical simplification (note the gut), enlargement of the nasarium, and reduction in size (the numbers show total lengths in mm). Illustrations from Stümpke (1961).
The forms that we’ve looked at so far belong to what Stümpke (1961) referred to as the monorrhinan or uni-snouter and asclerorrhinan or soft-nosed snouter divisions. Among the soft-nosed snouters, the most familiar and best known are the hopsorrhinidans or sclerorrhines: the snout leapers and kin. Equally well known are the members of the last division: the polyrrhinans, or multi-snouters. In the classic phyllogram produced by Bromeante de Burlas (1950), all major snouter divisions were posited as having originated from a common ancestor in a ‘hub and spokes’ type arrangement. This might have merit, especially given models that propose an explosive island-endemic radiation of rhinogradentians from a founding ancestor. However, the presence of fossil and extant rhinogradentians in the Old World has shown that all of the major divergences within Rhinogradentia had occurred during the Mesozoic, making the group an ancient one, with a relict distribution in modern times. This argues against the idea of an explosive island-endemic radiation.
The snout leapers or hopsorrhinidans. Snout leapers and their relatives included numerous terrestrial, arboreal and even volant forms. The earliest-diverging lineages within the group the arboreal snout leapers or perihopsids, likened by Stümpke (1961) to arboreally modified kin of Archirrhinos. Their overall proportions were not extraordinary, but their large nasarium was jointed like a limb and possessed a plate-like organ at its tip. The tail was similarly equipped, providing the animal with similar spring-action pseudo-limbs at both end of the body. Accordingly, perihopsids were reportedly extraordinarily agile, able to leap both forwards and backwards with speed (Stümpke 1961). Presumably derived from a perihopsid-like snout leaper, the hopsorrhinids or true snout leapers were characterised by an extraordinarily long, gracile, jointed nasarium that the animals used to make long backward leaps. The body was particularly short, hindlimbs were entirely absent, and the tail was long, slender and prehensile. Multiple species inhabited the beaches of Hi-yi-yi.

Caption: a hopsorrhinidan montage. Phyllohopla (upper right) was a mouse-sized, arboreal leaper, thought by Stümpke (1961) to be close to what ancestral forms of this group were like. Hopsorrhinids like the one shown at left had a far longer, more strongly jointed snout and were terrestrial. At lower right we see the Jurassic Otopteryx fossil (original image here), one of several fossils which demonstrates that most key divergences within rhinogradentians did not occur on island archipelagos in the Pacific.
The Earwing. The best known and most studied snout leaper – and indeed one of the best known and most studied of all rhinogradentians – was the Earwing Otopteryx volitans. While conventionally awarded its own taxonomic family it was more likely a derived hopsorrhinid as it certainly possessed all the derived peculiarities of this group. When terrestrial hopsorrhinids leapt, they were sometimes observed using their large ears to help steer their progression. This perhaps gives us a clue as to how the flight ability of Otopteryx evolved, as this remarkable rhinogradentian used its enormous wing-like ears in flight. The enormous aural cartilages and their hypertrophied muscles – particularly the m. aeroplano-jugalauris – allowed Otopteryx to flap its ears in rapid, skilful flight and it was even able to avoid avian predators on the archipelago (Stümpke 1961). Jurassic specimens of Otopteryx were reported from France in 1995 and are so similar to the Hi-yi-yi population that they were initially referred to the same species. This would be unprecedented for a mammal, and the possibility remains that the fossil should be named as a distinct taxon.

Caption: Earwing skeletal and muscular anatomy as depicted by Stümpke (1961). One detail that might not be apparent is that earwings flew tail-first, the jointed snout functioning as a steering tail held out behind the animal during flight. This might explain why the animal in Stümpke’s (1961) portrait – see below – is presenting its rump to us, not its face.
Orchidsnouters and lilysnouters. Finally among the snout leapers, there were the specialised orchidiopsids or orchidsnouters. These rhinogradentians are so unusual that their classification among snout leapers was initially not suspected (Stümpke 1961). Entirely arboreal, they lacked hindlimbs and hence probably descended from hopsorrhinids (in fact, it is probably accurate to regard them as part of Hopsorrhinidae, rather than as a separate family). Unlike hopsorrhinids, they possessed a large, flattened nasarium that resembled a leaf or petal. The large ears were also petal-like. Standing vertically on their powerful tail, and climbing slowly with their prehensile hands, they mimicked leaves and flowers and preyed on insects. At least one species, the Glowing lilysnouter Liliopsis thaumatonasus, was bioluminescent, this apparently being due to symbiotic bacteria (Stümpke 1961).

Caption: the most highly modified of the hopsorrhinidans: an Earwing at left, and an Ankel’s vanilla-perfumed orchidsnouter Orchidiopsis rapax at right. The Earwing was said by Stümpke (1961) to be beautifully coloured and iridescent, while orchidsnouters were attractively patterned and coloured, the vanilla-like aroma of this species being used to attract insects. A footnote in Stümpke (1961) noted that orchids were absent from the Hi-yi-yi islands, the actual flowers morphologically closest to orchidsnouter appendages being “closely related to Ranunculaceae” (that’s the buttercup-crowfoot group). Illustrations from Stümpke (1961).
The multi-snouters or polyrrhinans. Polyrhiny – the condition in which the nasarium is divided into several or multiple segments – was unique among rhinogradentians to the polyrrhinans, or multi-snouters. These are the most familiar of rhinogradentians, but also the most mysterious, as their origins remain obscure and they so differed from other rhinogradentians that they are generally thought to have diverged as early as the Cretaceous (Stümpke 1961). This has been validated by the discovery of hopsorrhinids in the Jurassic, as noted above.
Polyrrhinans were divided by Stümpke (1961) into the tetrarrhinidans (four-snouters), hexarrhinidans (six-snouters) and dolichoproatans (tasselsnouters). Tetrarrhinidans include the well-studied snout walkers or nasobemes and the amazing Predacious snouter Tyrannonasus imperator. The frugivorous nasobemes were able to walk on their four long snouts thanks to an internal skeleton of corpora spongiosa and a hydraulic system that provided turgidity when needed. The nasobeme tail was equally unusual: it was distally hollow and boneless and connected to the caecum. By forcing intestinal gas into the tail, a nasobeme could shoot out the ribbon-like segment for a distance of up to 3 m (this in a rhinogradentian with a nasarium and body length of about 1 m: they were among the largest members of the group). Tyrannonasus preyed preyed upon nasobemes, capturing them following pursuit on-nose and killing them with a venomous tail spike (Stümpke 1961). Various characters of the digestive tract indicate that Tyrannonasus evolved from frugivorous species and it has even been proposed that its predatory lifestyle developed from an original habit of stealing fruit from nasobemes, this piracy developing over time into predation.

Caption: at left, a Predacious snouter attacks a Nasobema, image from Stümpke (1961). At right, another of Takeshi Tokiwa’s remarkable models. Several images of this particular model are available online.
The six-snouters. Among the most specialised rhinogradentians, the hexarrhinidans (six-snouters) include the second group known as ribbon snouters, in addition to the six-flowered snouters and flower-faced snouters. Hexarrhinidan ribbon snouters (Eledonopsis) lived in burrows and were shrew-like, but for the ribbon-like multi-part nasarium that extended along the ground, some distance (up to c. 30 cm) away from the burrow. A ciliated epithelium extended along the entire dorsal surface of each nasarium segment, and small insects that became stuck here were carried by the cilia toward the mouth. Six-flowered snouters (Hexanthus) presumably descended from Eledonopsis-like forms but, instead of long ribbons, their proboscides took the form of long tubes, each sprouting a flower-like tip that the animal would wind about the stems of adjacent plants. Flying insects, deceived by these flower-like structures, would settle on them and be caught, and then moved by peristalsis along the tubes (Stümpke 1961).

Caption: at left, Eldenopsis suavis using its multi-branched nasarium to locate and collect arthropod prey. This was one of five known ribbon snouter species. At right, a vegetation stand featuring different ontogenetic stages of the remarkable flower-faced Cephalanthus. Illustrations from Stümpke (1961).
An apparent deviation from the cryptic lifestyle of hexarrhinidans was adopted by the flower-faced snouters (Cephalanthus). Equipped with petal-like fleshy growths surrounding the mouth, the best known members of this group remained sessile as remarkable flower-mimics. While young Cephalanthus had a ‘normal’ mammalian tail, in adults it was a long, stiff structure rooted in the ground. After birth, a young Cephalanthus would climb to the top of a tall flower, bite off the flower head, and then take up life as a flower-mimic, an exudate from the mouth attracting flying insects. The sessile attraction of insects, aided by oral exudates, may not be unique to rhinogradentians among tetrapods, as it has been suggested that frogmouths may also employ this technique (Diamond 1985). Cephalanthus is also strikingly similar to the sedentary Flooer Florifacies mirabila of the Batavian forests (Dixon 1981).
![Caption: the evolution within snouters of species over 1 m long means that they cannot be universally regarded as ‘small’ mammals. Mammontops was a mountain-dwelling herbivore, described by Stümpke as “relatively majestic”. The correct spelling of the name, incidentally, is indeed Mammontops, but it was accidentally written ‘Mamontops’ [sic] in the accompanying plate of Stümpke (1961).](https://images.squarespace-cdn.com/content/v1/510be2c1e4b0b9ef3923f158/1633693210760-PNU2OKAXXWMBFG5MUUW2/snouter-review-Oct-2021-Mamontops-alive-650px-100kb-Oct-2021-Darren-Naish-Tetrapod-Zoology.JPG)
Caption: the evolution within snouters of species over 1 m long means that they cannot be universally regarded as ‘small’ mammals. Mammontops was a mountain-dwelling herbivore, described by Stümpke as “relatively majestic”. The correct spelling of the name, incidentally, is indeed Mammontops, but it was accidentally written ‘Mamontops’ [sic] in the accompanying plate of Stümpke (1961).
While some of the polyrrhinans, such as the nasobemes and Predacious snouter, were quite large, even they were out-weighed by the Bearlike shaggyfaced snouter Mammontops ursulus, a herbivorous six-snouted species that attained a vertical height of 1.2 m in the males. Equipped with a thick woolly coat and short limbs that were well tucked-in to the pelage, it was a highland specialist that lived in small herds. Tasselsnouters, the dolichoproatans, were also large and possessed the most elaborate nasarial morphology of any rhinogradentian, with a ridiculous 18 snout pairs (Stümpke 1961).

Caption: an excellent montage which shows how large the largest of the snouters actually were. Here, we see (1) Shaggyfaced snouter and (2) Giant tasselsnouter to scale with Pioneer Dork, (3) Dixonian Nightstalker and (4) Black carpo (of the Speculative Dinosaur Project). Image: Fabio Alejandro.
Rhinogradentians alive! When Stümpke (1961) was published, rhinogradentians were regarded as unique to the Hi-yi-yi archipelago. As discussed earlier, however, Jurassic fossils show that snout leapers – members of one of the most recently evolved rhinogradentian clades – were formerly present in what is now continental Europe, suggesting that rhinogradentians were originally of very wide distribution. Several even more exciting recent discoveries have shown, amazingly, that relict rhinogradentians have survived to the present in various Old World locations. In 1999 a live rhinogradentian was discovered in Slovenia. Representing the new taxon Acrorrhinos, it appears to be particularly archaic and closely related to Haeckel’s primitive snouter. The discovery of this taxon is discussed here at Les Rhinogrades.

Caption: a discussion of how the living Slovenian snouter was discovered and photographed in 1999, as discussed here. The only photo has been reproduced at small size and very low resolution; sorry for the grainy look.
Three additional extant species were reported from the Weddell Sea in 2000 (Feldoyants & Zelbstander 2000): Rhizoidonasus euphorbiformis, Larvanasus haleciformis and Nudirhinus medusiformis. All are tiny (c 20 mm long), colonial, cnidarian-like animals, apparently part of the hexarrhinidan group, and demonstrate yet another ecomorphological variation within this group.
In 2004, another extant colonial rhinogradentian was discovered (Kashkina 2004), and later in the same year it was realised that various hermit crab parasites were also highly modified, tiny parasitic rhinogradentians (Bukashkina 2004). As discussed above, it was remarkable enough that the tiny turbellarian-like allsnouters were originally mistaken for invertebrates but, as ridiculous as it sounds, we now know of two snouter lineages that diverged from the mammalian body plan to such an unprecedented degree. It’s well known that a parasitic or infaunal lifestyle can lead to simplification in form and function, and these incredible tiny boneless mammals provide what might be the best example of this in the whole of zoology. Finally, three rather more ‘typical’ members of the group – the drillsnouters, all belonging to the genus Nasoperforator and noted for their wood-boring nasarium – were reported from the island of Espiritu Santo, Vanuatu, in 2012. They seem to have arrived on the islands via transportation in the wood of old ships!

Caption: at left, the tiny, ‘tree-nosed’, colonial snouter Dendronasus described by Kashkina (2004). It is aquatic, its highly modified forelimbs functioning as gills. At right, taxiderm specimen of Nasoperforator leguyaderi and illustration of N. bouffoni, both announced in 2012 by the Muséum National d’Histoire Naturelle.
Given that very little rhinogradentian material exists in collections, many species are known only from field sketches. Several workers have attempted in recent years to reconstruct some of the species three-dimensionally: Japanese model-maker Takeshi Tokiwa has produced models of many species (viewable here), as has Michaela Fortuber of the Naturhistorisches Museum at Braunschweig, Germany. Models are also on show at the Haus der Natur in Salzburg, Austria, and in a few other collections too. Naturally, rhinogradentians have made many appearances in the technical literature and their affinities and origins continue to be debated. It remains tragic that the vast majority of species are now extinct, especially when they were such unique, remarkable creatures.

Caption: assorted life reconstructions of snouters, variously observable online and even in a few museums of the world. At far, Takeshi Tokiwa’s model of a Golden toothed snout leaper. In the middle: especially cute nasobeme model. At far right: nasobeme model at the Haus der Natur, Salzburg. Images: Takeshi Tokiwa (far left); Haus der Natur (far right); I’ve been unable to find a credit for the image in the middle, let me know if you can help.
I’d like to thank everyone who has supported my academic interest in rhinogradentians over the years (particularly Dr Matt Wedel). And if it isn’t abundantly clear, the snouters are part of the SpecZoo canon. The article above was mostly written in-universe as per Stümpke’s (1961) original work.
For previous TetZoo articles on Speculative Zoology, see…
Dinosauroids revisited, November 2006
Dinosauroid cave art discovered, March 2007
How intelligent dinosaurs conquered the world, March 2008
Belatedly, Nemoramjetia (= Avisapiens), November 2008
Richard Dawkins and the crappy ‘humanoid dinosaurs’ that just won’t die, November 2009
Dinosauroids revisited, revisited, October 2012
Speculative Zoology, a Discussion, July 2018
Alternative Timeline Dinosaurs, the View From 2019 (Part 1), December 2019
Alternative Timeline Dinosaurs, the View From 2019 (Part 2), December 2019
Alternative Timeline Dinosaurs, the View From 2019 (Part 3): the Dinosauroid and its Chums, December 2019
Humanoid Dinosaurs Revisited Again: Russell and Séguin’s Dinosauroid at (Nearly) 40 Years Old, August 2021
Refs - -
Bromeante de Burlas, J. 1950. A derivacao e a arvore genealogica dos Rhinogradentes. Boll. Braz. Rhin. 2, 1203.
Bromeante de Burlas, J. 1952. The Hypogeonasidae. Bull. Darwin Inst. Hi. 5, Suppl.
Bukashkina, V. V. 2004. New parasitic species of colonial Rhinogradentia. Russian Journal of Marine Biology 30, 150.
Cain, J. 2018. In my tribe: what the snouters and jokelore reveal about tribes in science. Endeavour 43, 2-10.
Diamond, J. 1985. Filter-feeding on a grand scale. Nature 316, 679-680.
Dixon, D. 1981. After Man: A Zoology of the Future. Granada, London.
Feldoyants, S. D. & Zelbstander, A. 2000. Mysterious nose-walkers. Priroda 4, 46-54.
Kashkina, M. I. 2004. Dendronasus sp. – a new member of the order nose-walkers (Rhinogradentia). Russian Journal of Marine Biology 30, 148-150.
Stulten, D. 1955. The Evolution of Turbellarians, a Review of New Aspects. Piltdown University Press.
Stümpke, H. 1961. The Snouters: Form and Life of the Rhinogrades. The Natural History Press, Garden City, New York.



