Here we are with another effort to recycle material from the TetZoo archives (and invariably ruined by its hosters and hence made unavailable to those who might consult it). This time: a revamped version of my fossil pronghorns article, first published at ver 2 back in 2010 (an original is here)…

Caption: at left, the asymmetric Ramoceros (see below for more); at right, reconstructed Osbornoceros (though see below) with a colour scheme similar to – but different from – that of the extant Antilocapra. Images: Nobu Tamura CC BY-SA 4.0.
As we’ll see below, fossil pronghorns encompassed a good amount of diversity: there were kinds with deer-like pseudo-antlers as well as others that superficially resembled living African antelopes like kudu or nyala, there were tiny dwarf forms, and there were also some real oddballs, unlike anything before or since. All of this is well known among palaeomammalogists, but as usual the information isn’t widely available outside of the technical literature. My aim here is to provide a brief, introductory review to fossil pronghorns. For more in-depth treatments, the reader is directed to Janis & Manning (1997), Heffelfinger et al. (2004) and Davis (2007).
Pronghorns (or antilocaprids) are an entirely North American group. For reasons that aren’t clear, they never invaded South America when the Panamanian land-bridge formed nor, so far as we know, did they migrate into Asia. It might be that the general restriction of pronghorns to relatively arid habitats slowed or prevented their spread, but this doesn’t explain everything. During most of the Miocene, several pronghorn taxa were contemporaneous. This was true during the Middle Miocene when woodlands were widespread, but the largest radiation of the group coincided with the spread of savannah habitats in the Late Miocene. During the late Hemphillian North American Land Mammal Age (corresponding to the Late Miocene), as many as seven pronghorn genera were contemporaneous, some of which contained several species. All were advanced antilocaprine forms, often with complex horns.

Caption: the Capromeryx minor mount previously on show at the Page Museum, Los Angeles. C. minor has sometimes been called the Dwarf pronghorn and is famously associated with the La Brea tar pits. Other Capromeryx species are known from other Pleistocene locations. Image: Darren Naish.
‘Merycodontines’: moose-like palmation and extreme asymmetry
Several pronghorn taxa that are mostly restricted to the Early and Middle Miocene have historically been grouped together as the merycodontines*. These lack the characters that unite the antilocaprine pronghorns, and are generally small pronghorns with branched, sometimes antler-like, horns in the males. Females were hornless, a fact suggesting more solitary, more territorial habits than those of later, antilocaprine pronghorns. It’s generally agreed that ‘Merycodontinae’ is paraphyletic (Janis & Manning 1997), though note that (so far as I know) no detailed, comprehensive phylogenetic analysis of Antilocapridae has ever been published… I mean, other than Janis & Manning (1997). Since I published the first version of this article, I’ve become aware of Holley May Flora’s PhD work (completed in 2019) on pronghorn phylogeny. Flora found a somewhat different phylogeny from the one depicted below (based on that of Janis & Manning (1997)) but the approximate tree shape is similar.
* McKenna & Bell (1997) recognised a merycodontine group, but used the name Cosorycinae. Named by Cope in 1887, this has technical priority over Merycodontinae Matthew, 1904.
Among the ‘merycodontines’ is Merriamoceros coronatus from the late Early Miocene and early Middle Miocene of California, Nevada, Nebraska and Montana. Its short horns terminate in an expanded terminal portion that supports many short, finger-like tines: a palmate morphology superficially resembling that of moose.
Caption: at left, skull of Merriamoceras. At right: life reconstruction of M. coronatus by Nobu Tamura, CC BY-SA 4.0 (original here).
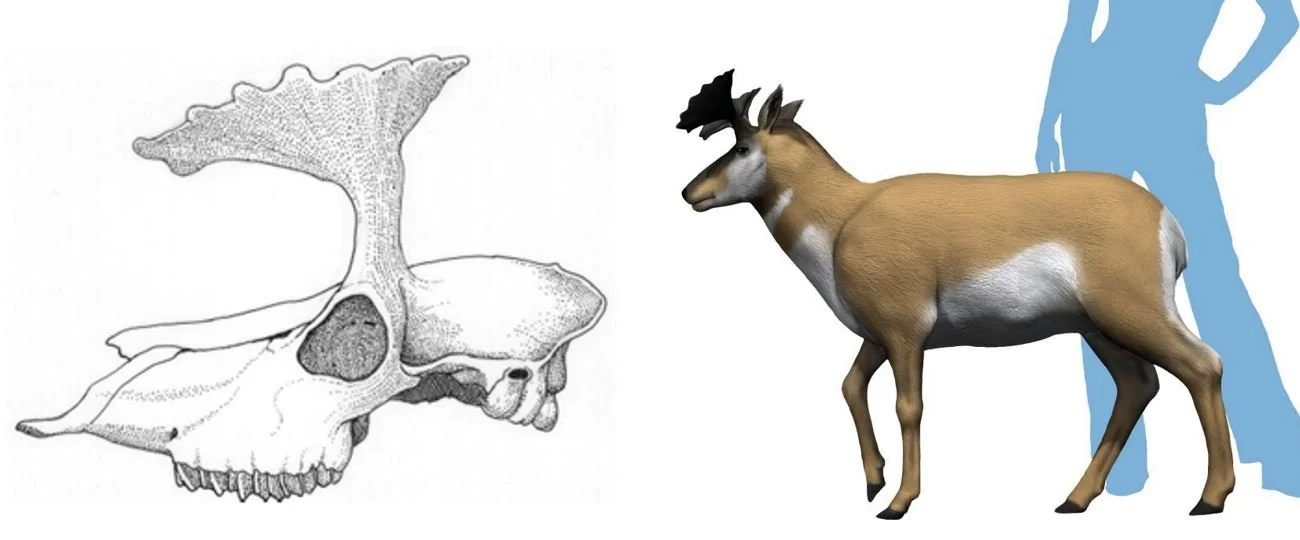
One of the most unusual pronghorns – in fact, one of the most unusual artiodactyls – also belongs to this region of the cladogram. I’m referring here to Ramoceros of the Middle and early Late Miocene. Several species (and even subspecies) have been named, and a second genus – Paramoceros – is nowadays included by most authors within Ramoceros. The main horn shaft is long, and three tines are present near the tip, forming a deer-like pseudo-antler. Oh, and – – one horn (it might be either the left or the right) is about twice the length of the other one. If the asymmetry was variable intraspecifically (and, so far as I can tell, it is), does this mean that ‘left handers’ (viz, those with a long horn on the left) could only fight with ‘right handers (viz, those with a long horn on the right), or what? I haven’t ever seen a discussion of this in the literature, but that doesn’t mean it doesn’t exist.
Caption: skull of Ramoceros ramosus, in posterior view, at left. At right: mounted skeleton of R. osborni on display at the American Museum of Natural History, New York. Images: Ryan Somma, CC BY-SA 2.0 (original here).
The surface texture of ‘merycodontine’ horns apparently indicates that they were skin-covered, rather than sheathed in horn. This is what it says in the literature, but keep in mind that it’s quite difficult to work out which tissues covered the horns of extinct mammals and there’s a historical tendency for people to claim the presence of a skin covering even when the evidence for it is not great.
Antilocaprine pronghorns: some bovid-like
The majority of pronghorns belong to Antilocaprinae, a clade characterised by a more posteriorly positioned orbit, reduction of the anterior tine on the horn and enlarged auditory bullae. The metapodials of antilocaprines are more elongate than those of other pronghorns, they all seem to have lacked the side toes that were present in more basal forms, and their premolars are proportionally larger and more molariform. All pronghorns have high crowned teeth; evidence from tooth wear indicates that Miocene species relied more on grass than does the living species (Semprebon & Rivals 2007). It might seem paradoxical that, while the living species has the highest crowned teeth of any pronghorn, only 12% or so of its diet is composed of grass. However, it’s been argued (e.g., Janis 1995) that it’s the soil and grit ingested with leaves – and not necessarily the abrasiveness of the leaves themselves – that controls hypsodonty in herbivorous mammals. It may therefore be that Antilocapra inhabits ‘dustier’ habitats than its fossil relatives.
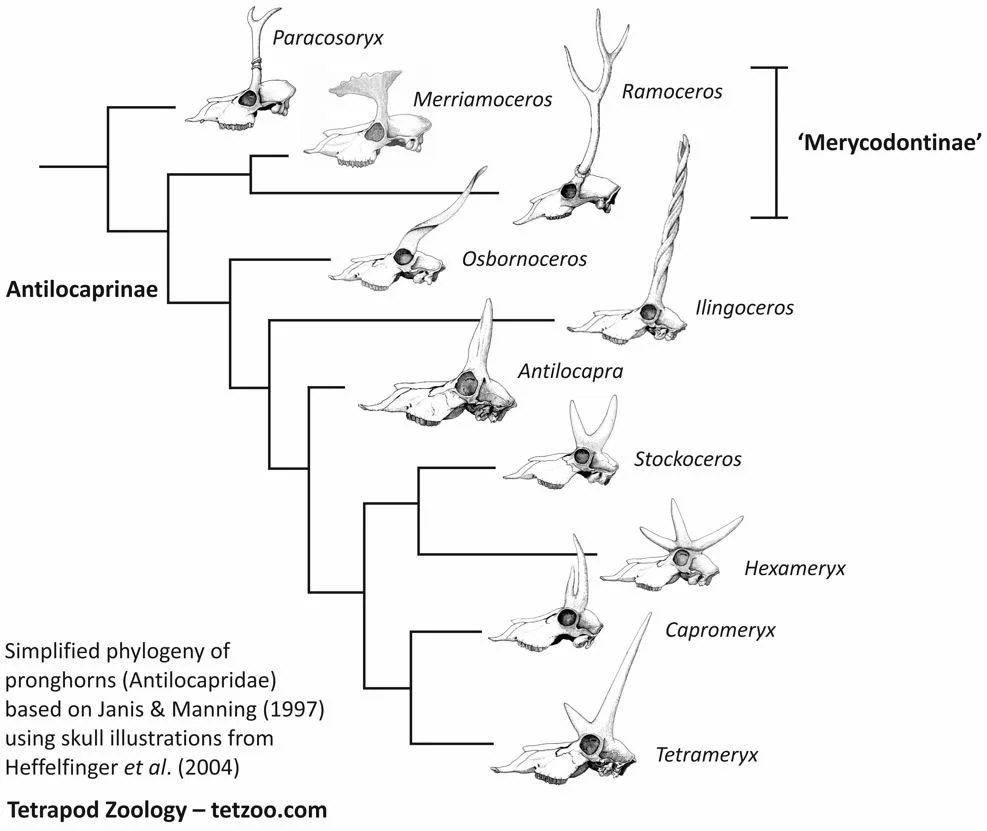
Most antilocaprine lineages possessed branched horns where a prominent anterior tine gave the bony horn-core a forked appearance. The anterior tine was, however, reduced or lost in some lineages and they then evolved long horns resembling those of gazelles or spiral-horned antelope. In Proantilocapra platycornea the anterior tine was absent and the horns were short, laterally compressed and with rounded tips. As suggested by its name, Proantilocapra was originally interpreted as close to the ancestry of Antilocapra (Barbour & Schultz 1934). The affinities of Antilocapra seem to lie elsewhere though. Some authors have identified Texoceros from the Late Miocene as its closest extinct relative (Janis & Manning 1997), whereas others find Stockoceros to be closest to Antilocapra… on which, read on.
Caption: skull of Proantilocapra platycornea in anterior and left lateral view, from Barbour & Schultz (1934), and life restoration labelled as Osbornoceros osborni (though this can’t be correct, since O. osborni has long, gently curved, gazelle-like horns). Image: Nobu Tamura CC BY-SA 4.0, from here.
The anterior tine was also lost in Osbornoceros osborni from the Late Miocene of New Mexico and Texas. This taxon possessed slender, posterolaterally twisted horns that look much like those of some gazelles or the Nyala Tragelaphus angasii.
Four-horned and six-horned antilocaprines
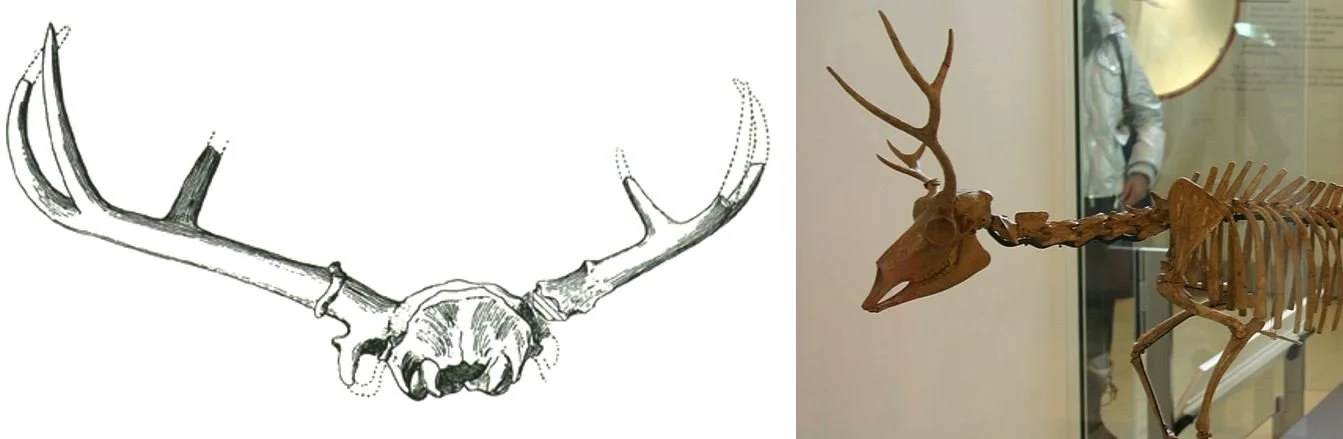
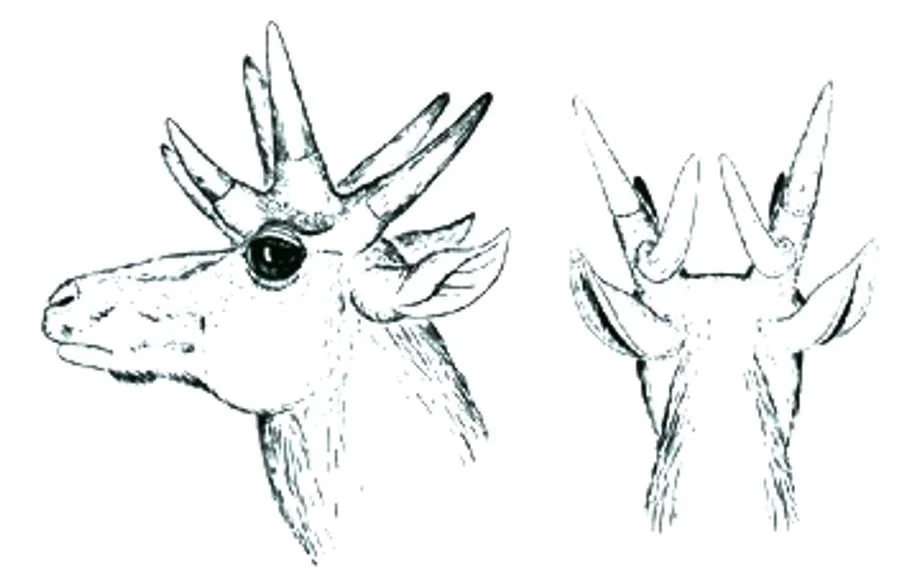
In some members of the antilocaprine clade Stockoceratini, the main shaft of the horn became reduced until it was virtually absent while, at the same time, the tines became enlarged. The result is that these pronghorns must have looked as if they had four (e.g., Hayoceros and Stockoceros) or even six (e.g., Hexameryx) horns, all emerging supraorbitally in a cluster. In the fossils of these animals, the horn tips have a different, smoother surface than the horn bases, suggesting that skin clothed the horn base and that keratin sheaths were restricted to the tips.

Caption: the Pleistocene pronghorn Stockoceros conklingi, known from the southern USA and Mexico, on display at the Los Angeles County Museum, LA. A second Stockoceros species (S. onusrosagris) is known from Arizona. Images: Darren Naish.
These enlarged tines are typically cylindrical in cross-section, but can be blade-like. To make things even weirder, in some stockoceratins (e.g., Hexobelomeryx), two of the three enlarged tines are so closely aligned that they must have been united in a single keratin sheath, though this was presumably forked like the underlying bone. Worth noting here is the fact that Antilocapra has unbranched bony horn-cores but a branching keratin sheath: could the unbranched horn-cores of fossil pronghorns have supported branched keratin sheaths? We really don’t know, and this possibility means that the reconstructions you see here – like Webb’s rendition of Hexameryx below – may well be conservative.
Caption: Webb’s (1973) reconstruction of Hexameryx simpsoni from the Pliocene of Florida.
Another particularly remarkable group of pronghorns are the ilingoceratins (their formal name is Ilingoceratini). As in Proantilocapra and Osbornoceros, the tines became reduced in this group, and in the most ‘extreme’ ilingoceratins the main shaft of the horn twists laterally. This is most prominently developed in Ilingoceros where two spiral ridges extend along the length of each horn such that they resemble the spiralling horns of kudus and other such antelopes. On naming I. alexandrae (two species of Ilingoceros are currently recognised), Merriam (1909) considered it to be what we would today call a tragelaphine bovid (the group that includes nyalas, kudus and elands). He later realised that it was an antilocaprid thanks to its high crowned teeth and other features. The horns in at least some Ilingoceros specimens have forked tips, but the forks don’t seem to be homologous to the tines present elsewhere among pronghorns.
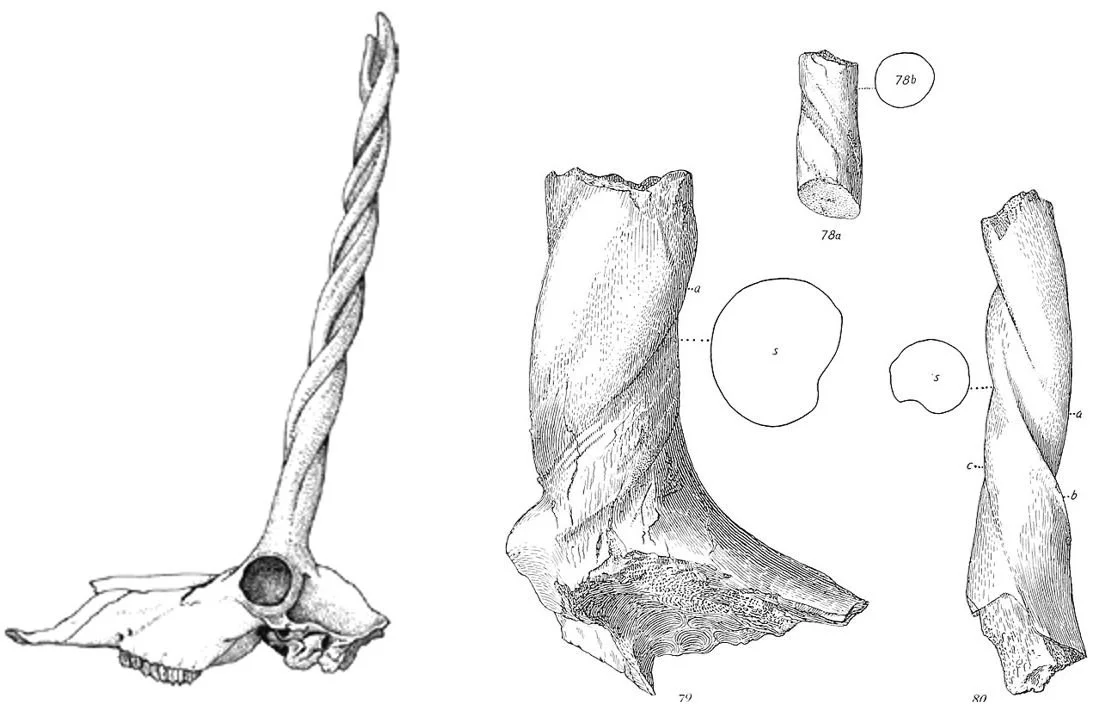
Caption: reconstructed skull of Ilingoceros, from Heffelfinger et al. (2004). At right, one of Merriam’s original Ilingoceros illustrations from his 1909 description of this animal.
I said earlier that some extinct pronghorns were tiny. Capromeryx of the Pliocene and Pleistocene – again, several species are known, and subspecies have even been named for some of them – first appears in the fossil record as an animal similar in size to the living Pronghorn (or, about 85% of it anyway). It then seems to have undergone a dwarfing trend, such that a species from the Late Pleistocene (namely C. minor, originally described from Rancho La Brea) was less than 60 cm tall at the shoulder. It might be presumed that these tiny pronghorns were secretive animals that lived like modern dik-diks or dwarf antelopes and were not denizens of open-habitats like their larger cousins. But their remains are in fact found in what were once grassland habitats, albeit in places where there were clumps of trees.

Caption: Capromeryx runs past the legs of the giant camel Titanotylopus. Image by Carl Buell, used with permission.
And so ends our brief look at the diversity of fossil pronghorns. I hope you enjoyed it! More on pronghorns later – there is still much to look at.
For previous TetZoo articles on pronghorns and other artiodactyls, see…
Duiker, rhymes with biker, August 2008
The monster sheep that wasn’t, and other tales of African Bovini, May 2011
Reasons for really liking wildebeest, February 2014
The Pronghorn Antelope: Designed by Committee, March 2014
This blog benefits from your support. Thank you to those who help via patreon!
Refs – –
Barbour, E. H. & Schultz, C. B. 1934. A new antilocaprid and a new cervid from the late Tertiary of Nebraska. American Museum Novitates 734, 1-4.
Davis, E. B. 2007. Family Antilocapridae. In Prothero, D. R. & Foss, S. E. (ed) Evolution of Artiodactyls. Johns Hopkins University Press, pp. 227-240.
Heffelfinger, J. R., O’Gara, B. W., Janis, C. M. & Babb, R. 2004. A bestiary of ancestral antilocaprids. Proceedings of the 20th Biennial Pronghorn Workshop 20, 87-111.
Janis, C. M. 1995. Correlations between craniodental morphology and feeding behavior in ungulates: reciprocal illumination between living and fossil taxa. In Thomason, J. (ed) Functional Morphology in Vertebrate Paleontology. Cambridge University Press, pp. 76-98.
Janis, C. M. & Manning, E. 1998. Antilocapridae. In Janis, C. M., Scott, K. M. & Jacobs, L. L. (eds) Evolution of Tertiary Mammals of North America. Volume 1: Terrestrial Carnivores, Ungulates, and Ungulatelike Mammals. Cambridge University Press, pp. 491-507.
McKenna, M. C. & Bell, S. K. 1997. Classification of Mammals: Above the Species Level. Columbia University Press (New York).
Merriam, J. C. 1909. The occurrence of strepsicerine antelopes in the Tertiary of northwestern Nevada. University of California Publications, Bulletin of the Department of Geology 5, 319-330.
Semprebon, G. M. & Rivals, F. 2007. Was grass more prevalent in the pronghorn past? An assessment of the dietary adaptations of Miocene to Recent Antilocapridae (Mammalia: Artiodactyla). Palaeogeography, Palaeoclimatology, Palaeoecology 253, 332-347.
Webb, S. 1973. Pliocene Pronghorns of Florida. Journal of Mammalogy 54



