Could it be true? Two new predatory dinosaurs from the UK – both spinosaurids, no less – described and named in the same paper?? Well…

Caption: life reconstruction of Ceratosuchops (at left) and Riparovenator, shown sharing a flooded depression on the Wessex Formation floodplain. Image: Anthony Hutchings.
The 1986 naming of the spinosaurid dinosaur Baryonyx walkeri remains one of the late 20th century’s most remarkable and newsworthy dinosaur-themed discoveries. Here was demonstration that somewhere as mundane and well-understood as southern England could still yield exciting new dinosaur species. Furthermore, Baryonyx is from the Wealden, a sedimentary unit that’s been combed for its fossils since the early 1800s at least. It’s literally one of the most intensively studied, thoroughly examined fossil-bearing parts of the geological column on the planet.
Within the Wealden, the original Baryonyx specimen is from the Upper Weald Clay Formation, a subdivision high up in the Wealden, dating to the Barremian part of the Early Cretaceous. As I always say in my Wealden-themed articles, Wealden stratigraphy and terminology is complex, and you’ll probably need to keep referring back to the simplified stratigraphic diagram I show here to work out what’s going on.
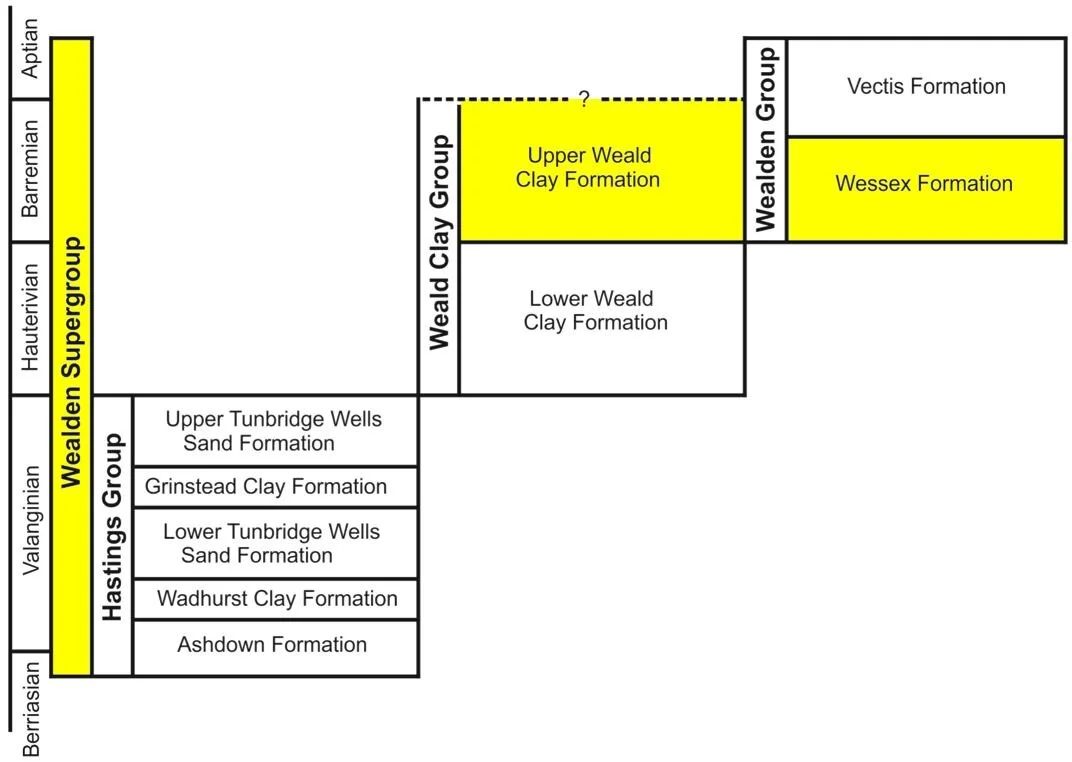
Caption: the simplified stratigraphic terminology for the Wealden that I’ve used on several occasions now. The Wealden as a whole is what’s known as a Supergroup, and it contains three formal Groups. The old Hastings Group was deposited in the Weald Sub-basin, as was the younger Weald Clay Group. The Wealden Group was deposited in the Wessex Sub-basin. Some of the sections more important to the article here are marked in colour.
The discovery of Baryonyx meant that here – at last – was our first good look at the appearance of the spinosaurids, a crocodile-faced group of theropods otherwise famous for the sail-backed, gigantic Spinosaurus of northern Africa (Charig & Milner 1986, 1990, 1997, Buffetaut 1989). By the time Baryonyx was monographed by the late Alan Charig and Angela Milner in 1997, it was obvious that spinosaurids of Baryonyx-like sort – baryonychines – were present throughout the whole of the Wealden, it’s just that their fragmentary fossils (namely, isolated teeth) had (understandably) been misidentified as those of crocodyliforms (Martill & Hutt 1996, Charig & Milner 1997, Naish et al. 2001, Buffetaut 2007, 2010, Naish 2011). Many had previously been labelled Suchosaurus, a name originally established by Richard Owen during the 1840s (Buffetaut 2007, 2010).

Caption: at left, the front page of Charig & Milner’s (1986) initial descriptive paper on Baryonyx. At right, the skeletal reconstruction and skull reconstruction provided in the 1997 monograph (Charig & Milner 1997). For more on the story of Baryonyx’s discovery, see this recent TetZoo article.
In the years since 1997, other baryonychine bits and pieces have been reported from the Wealden. They’re mostly teeth too but they also include an interesting tall-spined dorsal vertebra from the Isle of Wight’s Wessex Formation, a subdivision of the Wealden that includes some of its youngest sediments. European baryonychine remains have also been reported from the Lower Cretaceous of Spain (Ruiz-Omeñaca et al. 1997, 2005, Viera & Torres 1995) and Portugal (Buffetaut 2007, Mateus et al. 2011).
A widespread assumption has been that all of these remains can be assigned to Baryonyx, if not specifically to Baryonyx walkeri. While, in cases, this might be correct, there are reasons for thinking that it might very well be incorrect in some other cases. Some of these fossils are more than 10 million years older than the type specimen of Baryonyx walkeri. The rest of the Mesozoic record shows us that dinosaur species and genera generally lasted, at most, for one or two million years. Ergo, at least some of these animals almost certainly represent new taxa: not Baryonyx walkeri, and likely not Baryonyx at all. In addition, quite a few of these baryonychine fossils come from sedimentary settings and geographical locations distinct from the Upper Weald Clay Formation, with distinct dinosaur assemblages. I’ve therefore argued that baryonychine remains reported from outside the Upper Weald Clay Formation should be identified as cf. Baryonyx, Baryonyx sp. or Baryonyx cf. walkeri (Naish & Martill 2007, Naish 2011) (all of these designations mean slightly different things; let me know if you want elaboration). Furthermore, many of these fossils differ in detail from the remains of B. walkeri: many isolated teeth from the Wealden – including those labelled Suchosaurus – have flattened longitudinal strips on both of their sides, instead of just the lingual (inner) side, as is typical of the B. walkeri holotype.
Enough preamble – the main event… With this as our required background, we come at last to the main event. Today sees the publication of the new open access Scientific Reports paper ‘New spinosaurids from the Wessex Formation (Early Cretaceous, UK) and the European origins of Spinosauridae’, authored by Chris Barker, Dave Hone, myself and a team of colleagues (Barker et al. 2021). Therein, we preliminarily report two new baryonychine specimens from the Isle of Wight, both of which we interpret as new taxa.

Caption: known skeletal elements of Ceratosuchops (grey silhouette) and Riparovenator (black silhouette) to scale (scale bar = 100 cm). Image by Dan Folkes, from Barker et al. (2021).
Our new animals were discovered between 2013 and 2017, at or near beach level at Chilton Chine on the Isle of Wight’s south-west coast. One (represented by a braincase, snout-tip, additional skull fragments and an articulated partial tail) was discovered in situ in the sediments of the Wessex Formation. The other (represented also by a braincase and snout tip, and by other skull elements too) was found in loose sandstone blocks discovered on the foreshore. Because the stratigraphic position of the second specimen is uncertain, there’s no knowing how close it is in precise stratigraphic terms (and hence age) to the first. The fossils, incidentally, are curated in the collections of IWCMS, the Dinosaur Isle Museum (Isle of Wight County Museum Service) at Sandown, Isle of Wight.

Caption: Dinosaur Isle Museum at Sandown, Isle of Wight; the repository for the fossils discussed in this article. Image: Darren Naish.
Both animals differ in numerous ways from Baryonyx walkeri, both possess diagnostic features of their own, and both group away from Baryonyx walkeri on the cladogram and, surprisingly, on the same lineage as the baryonychine Suchomimus tenerensis* from Niger (Barker et al. 2021). Discovering two baryonychines that are both represented by three-dimensional snout-tip and braincase material is nothing short of amazing. We had both specimens scanned and printed (by Replicate 3D) for easy handling and comparison.
* A primary assumption of our taxonomic conclusions is that Suchomimus is recognised as a distinct genus relative to Baryonyx. The view that both should be synonymised has been mentioned on occasion but we follow the argument that they’re sufficiently distinct to warrant separation.

Caption: Suchomimus – shown here – is conventionally thought to have been longer-snouted than Baryonyx. The same might have been true of the two new taxa discussed here: they are more like Suchomimus than Baryonyx in numerous anatomical details. Credit: Hello!mrsax, CC BY-SA 4.0 (original here).
Two new baryonychines. It was immediately obvious that both Isle of Wight specimens differed in numerous ways. Lest anyone think that we were swift or lazy in our decision to name them as new taxa, let me emphasise that we went back and forth on what to do with the two repeatedly, and our final decision was not taken lightly. As stated in the paper, we remain open to the possibility that both might represent growth stages of the same taxon: the problem is that we lack appropriately comprehensive information on the changes that spinosaurids underwent as they matured (Barker et al. 2021). Furthermore, the features that make both different from each other and from Baryonyx don’t look like the sorts of changes that would occur during growth and exhibit a degree of variation that exceeds that seen within other theropod genera. Also leaning against the growth stage idea is that both specimens are similar in size, similar in size to the Baryonyx holotype as well, and exhibit similar degrees of skeletal fusion.
The better represented of the two – the one found stratigraphically in situ – is Riparovenator milnerae. The generic name means ‘riverbank hunter’ while the species name honours the late Dr Angela Milner, co-namer of Baryonyx. Angela’s recent passing coincidentally occurred while our work was underway and it seems absolutely right to have a baryonychine named after her.

Caption: braincases - in posterior view - of Ceratosuchops at left, and Riparovenator at right. The very different orientations of the paroccipital processes (pp) are obvious; other differences visible here include the differing locations of the subcondylar recesses (scr), the different shapes of the basisphenoids (bs) (they’re broad and flaring in Riparovenator but narrow and straight-edged in Ceratosuchops) and the presence of long, narrow basisphenoidal sulci in Ceratosuchops. Scale bars = 50 mm.
Riparovenator differs from other baryonychines in having a prominent notch in the bones that form part of its skull roof, an especially deep concavity high up on the back of the braincase, a distinctive placement of its facial nerve and in other cranial details too (Barker et al. 2021). The wing-like bony flanges that emerge from the rear surface of its braincase (termed paroccipital processes) are angled outwards and downwards more strongly in Riparovenator than they are in Baryonyx and in the second new animal… more on this in a moment. Finally, the tail vertebrae of Riparovenator are fascinating, their tall, narrow neural spines showing that the tail was deeper and more laterally compressed than thought before (they’re not, however, as extreme as the condition reportedly recently for Spinosaurus). We don’t say too much on the tail and its significance in this paper since we’re covering it in another study.
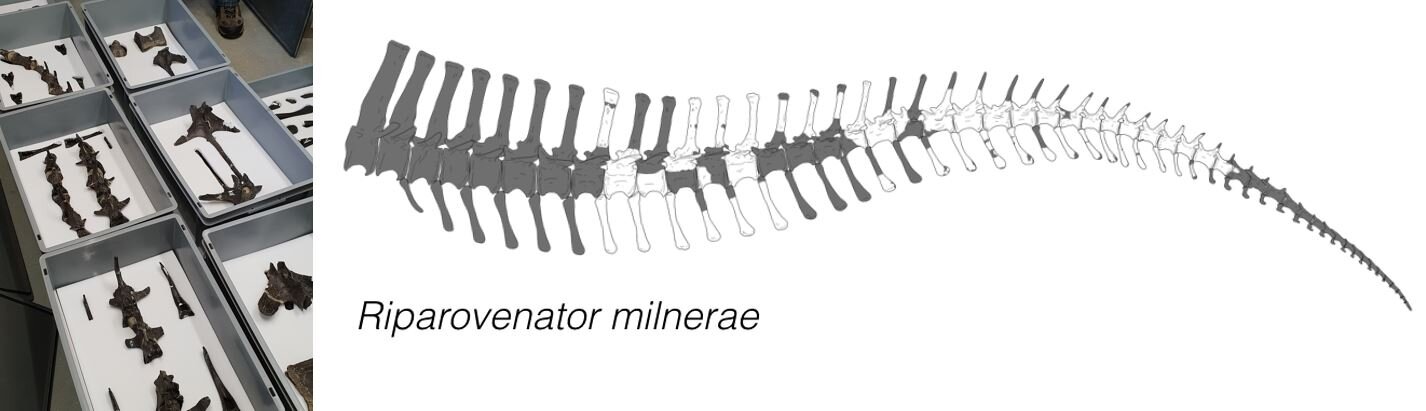
Caption: we’ve all been excited and intrigued by recently published data on the spinosaurine tail. Finally, we now have some idea of what the baryonychine tail is like too.
The second new taxon is Ceratosuchops inferodios, a name which roughly translates as ‘hell-heron horned crocodile-face’! Its key features include bony tuberosities on either side of the nostrils and long, deep bony hollows on the rear surface of the base of the braincase (Barker et al. 2021). As suggested by its name, Ceratosuchops also has a gnarly, elevated bony region around and above its eye (the bony bosses in this area were presumably elevated by horny coverings) as well as muscle attachment sites on the rear part of the skull that are bigger and more prominent than they are in other baryonychines (Barker et al. 2021).

Caption: at left, 3D prints of the skull roofs of Ceratosuchops (at top) and Riparovenator. Note the open notch in the orbital margin of Ceratosuchops (marked with arrow); the two are also quite different in the form and size of muscle attachments, bony crests and so on. At right: this 3D print of the Riparovenator braincase shows how good our prints are!
As mentioned above, a big surprise that became evident as we got to know these dinosaurs is that their anatomical features led them to group with Suchomimus, not Baryonyx, in phylogeny. We have thus opted to name a new clade within Baryonychinae: Ceratosuchopsini Barker et al., 2021.

Caption: reconstructed skulls of our two new taxa, shown on the assumption that their currently unknown sections were like those of Baryonyx. Both look similar but possess numerous features which were likely accentuated in life: Ceratosuchops, for example, has a lower midline premaxillary crest than other baryonychines and thus presumably had a ‘lower’ snout tip but was otherwise deeper and more muscular at the back of the skull. Images by Dan Folkes, from Barker et al. (2021).
Baryonychines in sympatry? So… three baryonychine taxa in the Wealden? Well, why not? For starters, Baryonyx walkeri, Ceratosuchops inferodios and Riparovenator milnerae are not demonstrably sympatric. They might all be Barremian in age, but the Barremian was four million years long and none of these dinosaurs are demonstrably from the same horizon. In addition, Baryonyx is from a different sedimentary basin from the one that yields Riparovenator and Ceratosuchops (Barker et al. 2021).
Moving on, the idea that Jurassic and Cretaceous dinosaur assemblages might contain two, three or more sympatric big theropods – even closely related ones – shouldn’t be considered abnormal. Conversely, it should be considered normal if we look at dinosaur assemblages worldwide. The Early/Middle Jurassic Cañadón Asfalto Formation of Argentina, the Late Jurassic Morrison Formation of the USA, the Late Cretaceous Judith River Formation of Canada – to name just a few – have also yielded several sympatric, similarly-sized, sometimes closely related big theropod taxa. This sort of thing was normal for the Mesozoic. Several previous studies have reported the sympatry of two or more spinosaurid taxa (e.g., Fanti et al. 2014, Samathi et al. 2021): this is not a first.

Caption: an especially picturesque view of Chilton Chine, discovery site of our two new dinosaurs. Image: Chris Barker.
Finally, at least some of the anatomical details that distinguish Baryonyx, Ceratosuchops and Riparovenator are of the sort that might well be linked to difference in lifestyle and ecology. Remember those diagnostic differences present at the back of the braincase in both new animals? These are exactly the sort of features that might be linked to different fishing styles, or differing abilities to grab, carry or process different sorts of prey. I put it that we’ve discovered different taxa that were doing different things: maybe some were more aquatically adapted than others, or better suited for tackling and lifting big prey than others, or better at grabbing small, swift-moving prey than others.

Caption: a reconstruction of Spinosaurus – with the fin-like tail and short hindlimbs but not the ‘M-shaped’ sail – produced for my new book Dinopedia. This individual is resting on a riverbank. Image: Darren Naish.
Some implications for Spinosaurus. On the subject of sympatry and taxonomic distinction… our paper also has relevance for a larger debate, and one considered almost inescapable among those researchers interested in Cretaceous theropods. I’m talking about the intractable mess that is Spinosaurus. As you’ll know if you’ve been following any of the post-2014 claims and counter-claims about north African spinosaurids, inherent to the Ibrahim et al. (2014) idea about Spinosaurus is that all the material in question belongs to the same one taxon.
In the interests of testing this idea, we coded several of the key specimens (including the Sigilmassasaurus holotype and ‘Spinosaurus B’) and included them in a phylogenetic analysis. The result? Well, things are ambiguous: in some versions of our analysis, the north African specimens did group together but the statistical support for this grouping was weak. Basically, it’s by no means clear that the specimens combined to make the chimaeric view of ‘Spinosaurus’ preferred by some authors really do belong to the same clade and same taxon. My personal view remains that the specimens concerned represent three or more taxa.

Caption: time-calibrated spinosaurid phylogeny from Barker et al. (2021). See the paper for attribution of silhouettes.
The other spinosaurids. What of other spinosaurids, and what of the larger picture? In general, we found support for the idea that Spinosauridae contains the two clades Baryonychinae and Spinosaurinae. Like a few other authors, however, we didn’t always get this result, since baryonychines were sometimes recovered as a paraphyletic grade to spinosaurines. Other authors have reported this result in the past (Evers et al. 2015, Sales & Schultze 2017). Among other interesting things, we recovered Vallibonavenatrix from Spain – described as a spinosaurine – as either a baryonychine or as outside the baryonychine + spinosaurine clade, while Camarillasaurus from Spain was found to be a spinosaurine (Barker et al. 2021). The Brazilian Irritator jumped around within Spinosaurinae, suggesting that data is needed on its postcranial skeleton (we coded only for the holotype skull) before we can better pin it down.
Out of Europe. Finally, when our phylogenetic framework for Spinosauridae was combined with data on the distribution of the taxa concerned, we found spinosaurids to be an ancestrally European group, not a Gondwanan or cosmopolitan one as sometimes argued in the past. There’s still a lot to learn – the almost total absence of Jurassic spinosaurid taxa remains a problem! – but, for now, it looks as if the group originated in Europe, then spread to Asia and Africa. The presence of both baryonychines and spinosaurines in Africa indicates separate migration events for these clades.

Caption: our study mostly supports the idea that Spinosauridae consists of the two clades Spinosaurinae and Baryonychinae, representatives of which are shown here. However, it remains possible that baryonychines are a grade, not a clade. Credit: Darren Naish.
That about wraps things up for now. I’ve been involved in work on these Wessex Formation baryonychines for as long as they’ve been known, and what was originally planned to ‘merely’ be the reporting of what were assumed to be additional Baryonyx specimens has ended up developing into something grander and more complex. More work on European and African baryonychines – and more detailed descriptive work on Riparovenator, Ceratosuchops and even Baryonyx itself – is underway and set to appear in the future.
And 2021 isn’t over yet. Will more Wealden-themed dinosaur news appear before the year is through? Stay tuned….
For previous TetZoo articles on spinosaurids, British theropods and associated issues (some links here are to wayback machine versions due to destruction or paywalling of everything at versions 2 and 3), see…
Of Becklespinax and Valdoraptor, October 2007
The world’s most amazing sauropod, November 2007
Oh no, not another new Wealden theropod!, June 2009
Concavenator: an incredible allosauroid with a weird sail (or hump)... and proto-feathers?, September 2010
The Wealden Bible: English Wealden Fossils, 2011, November 2011
Ostrich dinosaurs invade Europe! Or do they?, June 2014 (every archived version of this article lacks the original illustrations, sorry)
Theropod Dinosaurs of the English Wealden, Some Questions (Part 1), March 2020
Introducing ‘Unexpected Isle of Wight Air-Filled Hunter’, a New English Theropod Dinosaur, September 2020
Dr Angela Milner and the Discovery of Baryonyx, August 2021
You can support this blog – and my work in general – at patreon.
Refs - -
Buffetaut, E. 1989. New remains of the enigmatic dinosaur Spinosaurus from the Cretaceous of Morocco and the affinities between Spinosaurus and Baryonyx. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 1989, 79-87.
Buffetaut, E. 2007. The spinosaurid dinosaur Baryonyx (Saurischia, Theropoda) in the Early Cretaceous of Portugal. Geological Magazine 144. 1021-1025.
Buffetaut, E. 2010. Spinosaurs before Stromer: early finds of spinosaurid dinosaurs and their interpretation. In Moody, R. T. J., Buffetaut, E., Naish, D. & Martill, D. M. (eds) Dinosaurs and Other Extinct Saurians: A Historical Perspective. Geological Society, London, Special Publications 343, pp. 175-188.
Charig, A. J. & Milner, A. C. 1986. Baryonyx, a remarkable new theropod dinosaur. Nature 324, 359-361.
Charig, A. J. & Milner, A. C. 1990. The systematic position of Baryonyx walkeri in the light of Gauthier’s reclassification of the theropoda. In Carpenter, K. & Currie, P. J. (eds), Dinosaur Systematics: Approaches and Perspectives. Cambridge University Press, pp. 127-140.
Charig, A. J. & Milner, A. C. 1997. Baryonyx walkeri, a fish-eating dinosaur from the Wealden of Surrey. Bulletin of the Natural History Museum 53, 11-70.
Fanti, F., Cau, A., Martinelli, A. & Contessi, M. 2014. Integrating palaeoecology and morphology in theropod diversity estimation: a case from the Aptian-Albian of Tunisia. Palaeogeography, Palaeoclimatology, Palaeoecology. 410, 39-57.
Hutt, S. & Newbery, P. 2004. An exceptional theropod vertebra from the Wessex Formation (Lower Cretaceous) Isle of Wight, England. Proceedings of the Isle of Wight Natural History & Archaeology Society 20, 61-76.
Ibrahim, N., Sereno, P. C., Dal Sasso, C., Maganuco, S., Fabri, M., Martill, D. M., Zouhri, S., Myhrvold, N. & Lurino, D. A. 2014. Semiaquatic adaptations in a giant predatory dinosaur. Science 345, 1613-1616.
Martill, D. M. & Hutt, S. 1996. Possible baryonychid dinosaur teeth from the Wessex Formation (Lower Cretaceous, Barremian) of the Isle of Wight, England. Proceedings of the Geologists’ Association 197, 81-84.
Mateus, O., Araújo, R., Natário, C. & Castanhinha, R. 2011. A new specimen of the theropod dinosaur Baryonyx from the early Cretaceous of Portugal and taxonomic validity of Suchosaurus. Zootaxa 2827, 54-68.
Naish, D. 2011. Theropod dinosaurs. In Batten, D. J. (ed.) English Wealden Fossils. The Palaeontological Association (London), pp. 526-559.
Naish, D., Hutt, S. & Martill, D. M. 2001. Saurischian dinosaurs 2: Theropods. In Martill, D. M. & Naish, D. (eds) Dinosaurs of the Isle of Wight. The Palaeontological Association (London), pp. 242-309.
Naish, D. & Martill, D. M. 2007. Dinosaurs of Great Britain and the role of the Geological Society of London in their discovery: basal Dinosauria and Saurischia. Journal of the Geological Society, London 164, 493-510.
Ruiz-Omeñaca, J. I., Canudo, J. I., Cruzado-Caballero, P., Infante, P. & Moreno-Azanza, M. 2005. Baryonychine teeth (Theropoda: Spinosauridae) from the Lower Cretaceous of La Cantalera (Josa, NE Spain). Kaupia 14, 59-63.
Ruiz-Omeñaca, J. I., Canudo, J. I. & Cuenca-Bescós, G. 1997. First evidence of baryonychid dinosaurs (Saurischia: Theropoda) in the upper Barremian (Lower Cretaceous) of Vallipon (Castellote, Teruel, Spain). Beca del Museo de Mas de las Matas 17, 201-223.
Samathi, A., Sander, P. M. & Chanthasit, P. 2021. A spinosaurid from Thailand (Sao Khua Formation, Early Cretaceous) and a reassessment of Camarillasaurus cirugedae from the Early Cretaceous of Spain. Historical Biology doi: 10.1080/08912963.2021.1874372
Viera, L. I. & Torres, J. A. 1995. Presencai de Baryonyx walkeri (Saurischia, Theropoda) en el Weald de La Rioja (Espana). Nota previa. Munibe 47, 57-61.



