Time will tell. This is really exciting stuff.
If you enjoyed this article and would like to see me do more, please consider supporting this blog (for as little as $1 per month) at patreon. The more support I receive, the more financially viable this project becomes and the more time and effort I can spend on it. Thank you :)
For previous TetZoo articles relevant to this one, see…
Refs - -
Daza, J. D., Bauer, A. M., Stanley, E. L., Bolet, A., Dickson, B. & Losos, J. B. 2018. An enigmatic miniaturized and attenuate whole lizard from the mid-Cretaceous amber of Myanmar. Breviora 563, 1-18.
Daza, J. D., Stanley, E. L., Wagner, P., Bauer, A. M. & Grimaldi, D. A. 2016. Mid-Cretaceous amber fossils illuminate the past diversity of tropical lizards. Science Advances 2 (3), e1501080.
Lee, M. S. Y., Cau, A., Naish, D. & Dyke, G. J. 2014. Sustained miniaturization and anatomical innovation in the dinosaurian ancestors of birds. Science 345, 562-565.
Wang, M. & Zhou, Z. 2017. The evolution of birds with implications from new fossil evidences. In Maina, J. N. (ed) The Biology of the Avian Respiratory System. Springer International Publishing, pp. 1-26.
Xing, L., McKellar, R. C., O’Connor, J. K., Bai, M., Tseng, K. & Chiappe, L. M. 2019. A fully feathered enantiornithine foot and wing fragment preserved in mid-Cretaceous Burmese amber. Scientific Reports 9, 927.
Xing, L., McKellar, R. C., Wang, M., Bai, M., O’Connor, J. K., Benton, M. J., Zhang, J., Wang, Y., Tseng, K., Lockley, M. G., Li, G., Zhang, Z. & Xu, X. 2016a. Mummified precocial bird wings in mid-Cretaceous Burmese amber. Nature Communications 7, 12089.
Xing, L., McKellar, R. C., Xu, X., Li, G., Bai, M., Scott Persons IV, W., Miyashita, T., Benton, M. J., Zhang. J., Wolfe, A. P., Yi, Q., Tseng, K., Ran, H. & Currie, P. J. 2016b. A feathered dinosaur tail with primitive plumage trapped in mid-Cretaceous amber. Current Biology 26, 3352-3360.
Xing, L., O’Connor, J. K., McKellar, R. C., Chiappe, L. M., Bai, M., Tseng, K., Zhang, J., Yang, H., Fang, J. & Li, G. 2018. A flattened enantiornithine in mid-Cretaceous Burmese amber: morphology and preservation. Science Bulletin 63, 235-243.
Xing, L., O’Connor, J. K., McKellar, R. C., Chiappe, L. M., Tseng, K., Li, G. & Bai, M. 2017. A mid-Cretaceous enantiornithine (Aves) hatchling preserved in Burmese amber with unusual plumage. Gondwana Research 49, 264-277.
Xing, L., O’Connor, J. K., Schmitz, L., Chiappe, L. M. McKellar, R. C., Yi, Q. & Li, G. 2020. Hummingbird-sized dinosaur from the Cretaceous period of Myanmar. Nature 579, 245-249.





![It’s well known in the theropod research community that the full description of this amazing fossil - the holotype of the Spanish ornithomimosaur Pelecanimimus - was done back in the 1990s [UPDATE: nope, 2004], but hasn’t seen print for a bunch of r…](https://images.squarespace-cdn.com/content/v1/510be2c1e4b0b9ef3923f158/1585002681161-29JO3RG3SRL02I363HHH/Wealden-theropod-questions-Mar-2020-Pelecanimimus-holotype-skull-1274px-250kb-Mar-2020-Darren-Naish-Tetrapod-Zoology.JPG)






























































































































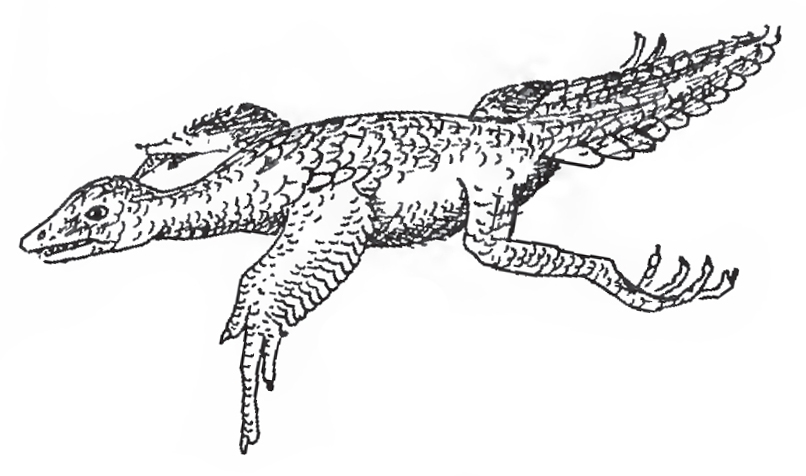
![William Beebe’s hypothetical tetrapteryx creature. A pre-Archaepteryx glider. You’ll note that Beebe was a pretty good artist [UPDATE: this is an error. The illustrations in this publication were by Dwight Franklin, not by Beebe!]. Image: Beebe (191…](https://images.squarespace-cdn.com/content/v1/510be2c1e4b0b9ef3923f158/1546356482204-OC8JSKSSMTH2YL41VQNL/Heilmann-Proavian-Beebe-Tetrapteryx-Jan-2019-Tetrapod-Zoology.jpg)