On Sunday 2nd September 2018, the immersive, interactive time-travelling visitor attraction known as Dinosaurs in the Wild closed its doors for the last time. Yes, Dinosaurs in the Wild is now officially off-show, and if you didn’t get to see it before that fateful Sunday… where were you? I was determined to embark on one final tour, and of course I also needed to go to the grand send-off party and say those sad final goodbyes…

Preparing to embark on a last ever tour of Late Maastrichtian western North America. Chrononaut Jasmine Arden-Brown introduces us to the world of Chronotex. Image: Darren Naish.
As discussed at Tet Zoo several times already (all at ver 3, I hasten to add), I was scientific advisor for this grand project and thus very much feel that the look, behaviour and biology of the Late Cretaceous animals brought to life for the experience was and is ‘mine’, the MAJOR disclaimers being (1) that a whole team of people actually did the work that resulted in the vision coming to fruition, and (2) any ideas that I have about extinct animal biology or appearance or whatever involve the proverbial standing on the shoulders of giants, and the work and discoveries of a great many other people.

As with any project of this size and scale, there's the long process whereby models and other props come together over time, and then there's the concept art, the explanatory diagrams, and so on. I've kept a record of as much of this stuff as I could. Image: Darren Naish.
With its fully – indeed, extensively – feathered dromaeosaurs, fuzzy-coated, muscular tyrannosaurs, terrestrial stalking azhdarchid pterosaurs (cough cough Witton & Naish 2008 cough cough), sleek, chunky mosasaurs, balloon-faced ankylosaurs and more (Conway et al. 2012), Dinosaurs in the Wild has – I really hope and feel – introduced a substantial chunk of the human public to a very up-to-date view of the Mesozoic world, and has thus gone some way towards undoing the damage of Jurassic World. No to the scaly, shit-brown, roaring monsters of the past, and yes to a more interesting, biologically plausible and often more surprising view of what these animals were like. Incidentally, Colin Trevorrow visited Dinosaurs in the Wild within the last few weeks, spoke to our associate live action director Cameron Wenn, and said really positive things (Colin and I spoke briefly over twitter).

Here are two of the (normally nocturnal) Dinosaurs in the Wild animals seen in full illumination. At left, the metatherian mammal Didelphodon; at right, the small dromaeosaur Acheroraptor (it never stays still for long, hence the motion blur). Image: Darren Naish.
And did Dinosaurs in the Wild have an impact on the public? I don’t know if I’m allowed to release all the figures, but I will say that many thousands of people attended the experience during its 13 or so months of operation at Birmingham, Manchester and London. Our amazing actors and other staff all became worthy ambassadors of ‘new look’ Mesozoic animals and their biology, and the substantial amount of scientific content included in the show surely introduced the public to a great deal of information they haven’t seen or heard before. All results indicate that we certainly received the sort of feedback and accolade we hoped for: we scored really well as goes visitor feedback, indeed sufficiently well that Dinosaurs in the Wild can be regarded as a world class attraction. The palaeontologists and other scientists and experts who visited were all extraordinarily positive, and thanks indeed to those colleagues of mine who voiced their thoughts in public (Dean Lomax, Mark Witton, Albert Chen, Dave Hone, among others).

Our final goodbye party was a solemn, quiet affair. Obviously. Thanks, Mike. Image: Darren Naish.
Our venues were all great – Manchester’s Event City was certainly quite the sight to behold – but were perhaps not as centrally placed as might be ideal, though there are all kinds of factors controlling where and how a given exhibit can be located.

A very dangerous box. Working in the Mesozoic is not all that easy. Image: Darren Naish.
Even now, and even after me writing that fairly substantial ‘behind the scenes’ article I published at Tet Zoo ver 3 in July 2018 (and here’s assuming that SciAm haven’t removed it due to an issue with image rights, ha ha ha), there’s a huge amount that could be said about the ‘making of’ this project. As some of you already know, the backstory to the world of Dinosaurs in the Wild is already written-up in an extensive document that we took to referring to as The Bible, but despite efforts I’ve had to give up on plans to get it published. I will be talking about much of the ‘behind the scenes’ stuff at TetZooCon this year (BUY TICKETS HERE), however, and will be bringing The Bible along for those interested in seeing it.

There's so much to see through the windows that, even after multiple visits, I still haven't seen it all. In this sequence (seen while looking across the Dakotaraptor nesting colony), two female dromaeosaurs engage in a squabble. Image: Kerry Mulvihill.
Huge thanks to everyone at the event last night, and to everyone who made Dinosaurs in the Wild the success it was. Special thanks to producers Jill Bryant and Bob Deere, creative director Tim Haines, the team at Freeman Ryan, to live action directors Scott Faris and Cameron Wenn, to all the amazing people at Impossible, Milk VFX and Crawley Creatures, to every single one of our amazing actors, to our support staff, our sponsors and everyone else. And thanks also to Sam, Simon, Heather and the others who accompanied me on the same, final tour I took just yesterday.

The temporal field generator is always on. Image: Darren Naish.
Is this really the very end of Dinosaurs in the Wild? Will it re-appear elsewhere at some future date? As you might guess, there are certainly plans that such a thing might happen. Right now, I can say no more. News will be reported here, and on relevant social media streams (I'm at @TetZoo, and check @DinosInTheWild too).
Refs - -



































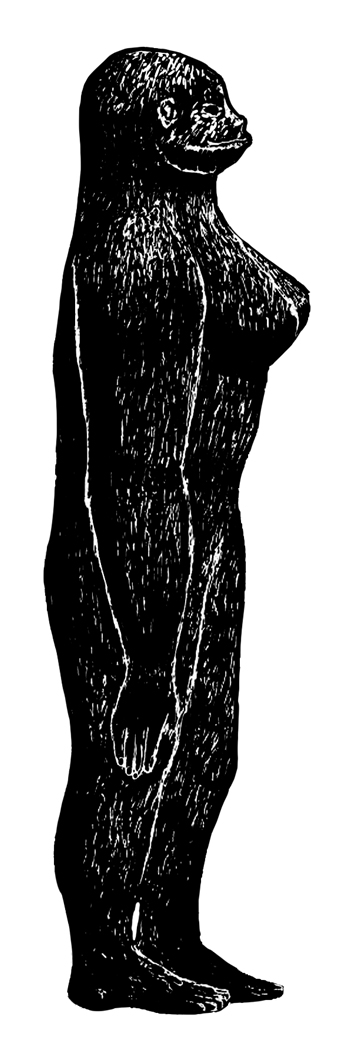
![I've only had one up-close encounter with a Bigfoot myself (it happened in California); I didn't have opportunity at the time to do any checking as goes any details of anatomy, but here's the proof. Image: [safely anonymous source]/Darren Naish.](https://images.squarespace-cdn.com/content/v1/510be2c1e4b0b9ef3923f158/1533161346909-MMK41T4JOD99AEY17TJ4/Bigfoot-in-LA-572-px-tiny-Aug-2018-Darren-Naish-Tetrapod-Zoology.jpg)